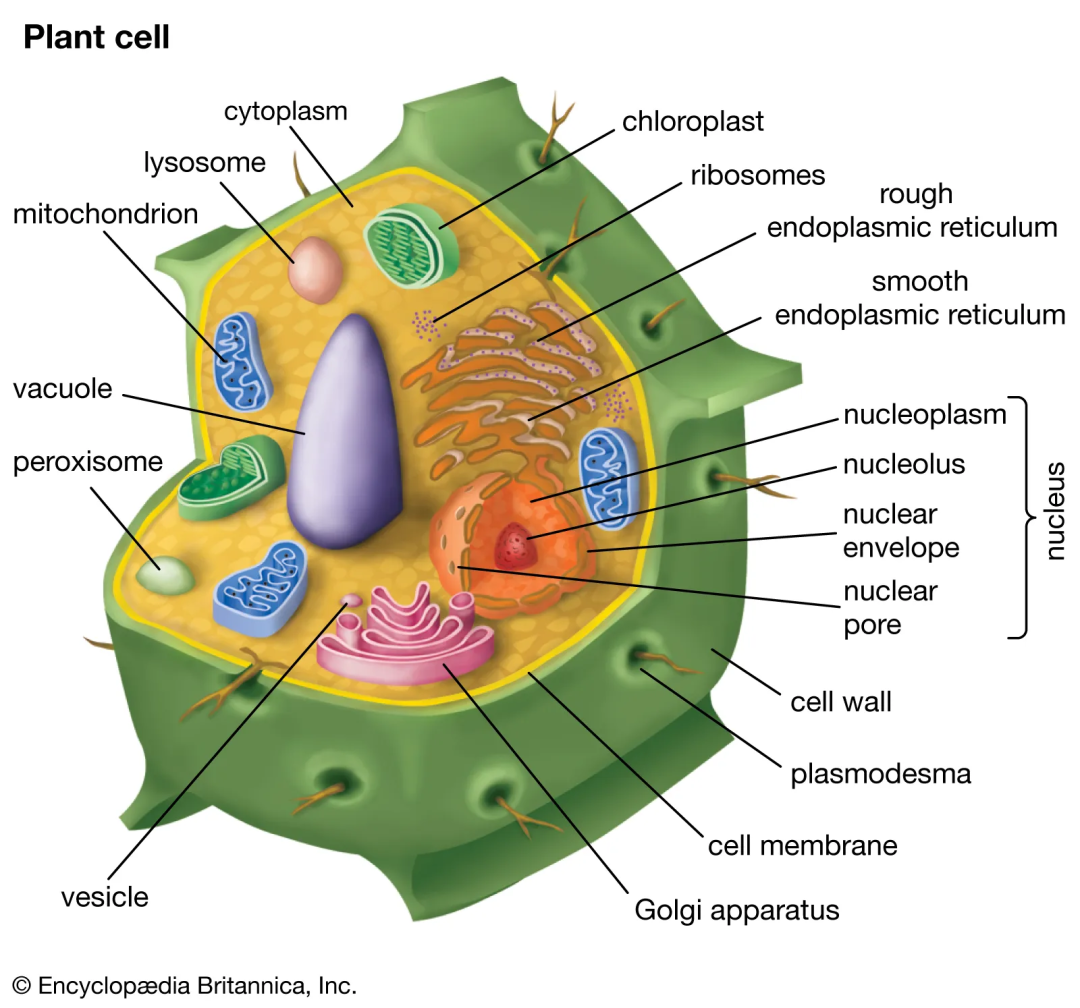
图1 植物亚细胞结构图,来源britannica。
亚细胞定位是指某种蛋白或表达产物在细胞内的具体存在部位,例如在核内、胞质内或者细胞膜上存在。在真核细胞中,除少量蛋白质在线粒体和叶绿体内合成外,绝大多数蛋白质都是由核基因编码,或在游离的核糖体上合成,或在糙面内质网膜结合的核糖体上合成。蛋白质合成以后必须转运到特定的部位才能参与组装细胞结构,发挥其生物学功能,这一过程称为蛋白质亚细胞定位。蛋白质的功能、代谢以及相互作用等都与其亚细胞定位密切相关,成熟蛋白质必须在特定的亚细胞结构中才能发挥正确的生物学功能,如果定位发生偏差,将对细胞功能甚至生命产生重大影响,因此对蛋白质亚细胞定位的研究具有重要意义。
研究亚细胞定位的方法:
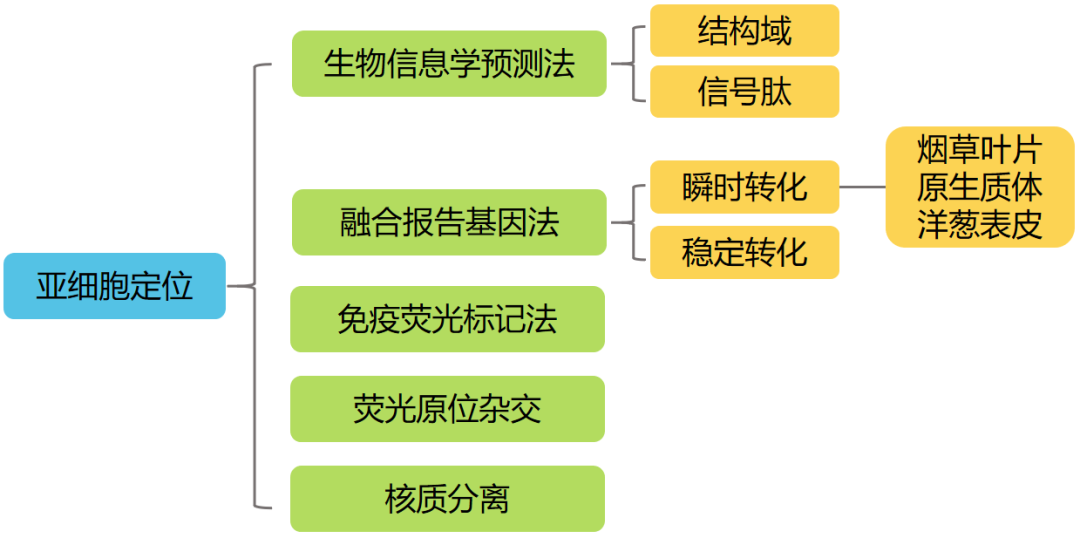
伯远生物亚细胞定位采用的方法主要是借助荧光蛋白报告基因来实现目标蛋白定位的融合报告基因定位法,原理主要是将目标蛋白与荧光蛋白的N端或者C端融合,通过瞬时转化技术,使得该融合蛋白在受体材料细胞内表达,目标蛋白会牵引荧光蛋白一起定位到目标细胞器,在扫描共聚焦显微镜的激光照射下会发出绿色荧光,从而可以精确地定位蛋白质的位置。融合报告基因定位法具有许多优势:适用性强,操作简便,周期较短,可用于活体的实时定位及动态研究,灵敏度高等。
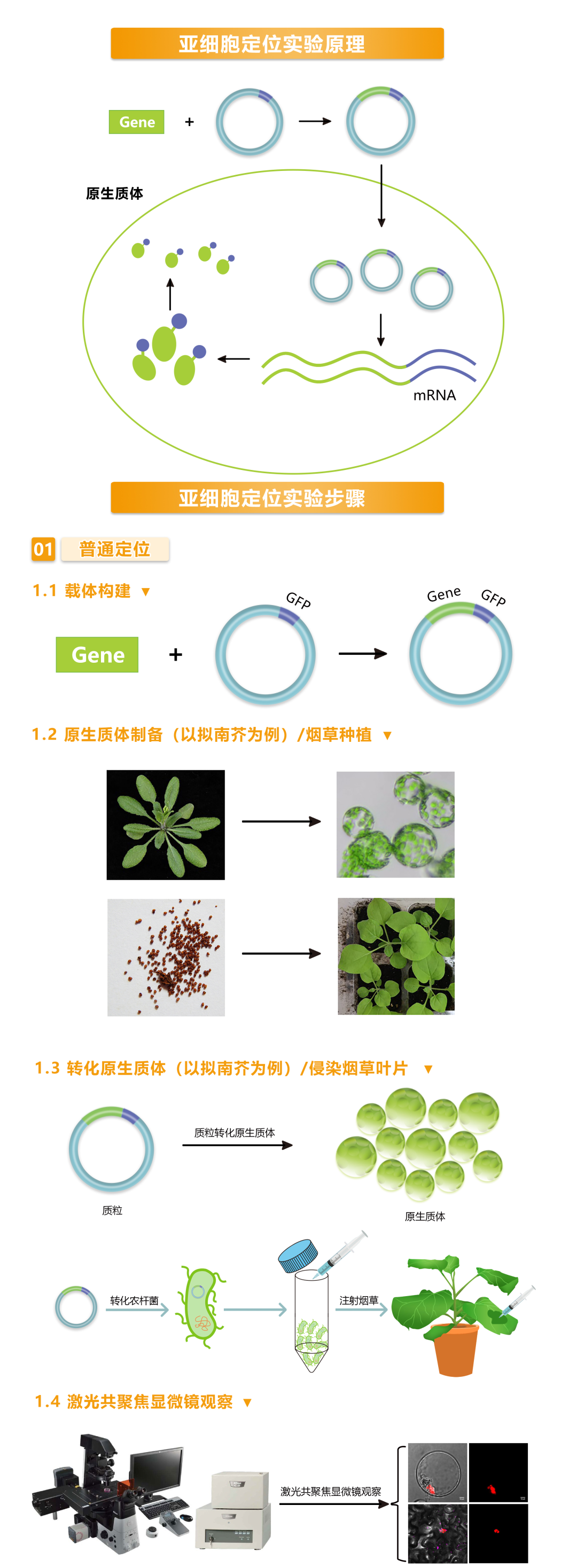
图2 伯远生物亚细胞定位流程。
之前有很多小伙伴咨询小远亚细胞定位与亚细胞共定位的区别,小远在这里给大家简单讲解一下,亚细胞共定位就是在普通亚细胞定位的基础上,同时转入一个Marker载体,以明确目标蛋白具体的定位信息,相比于普通亚细胞定位,共定位得到的结果更加准确。
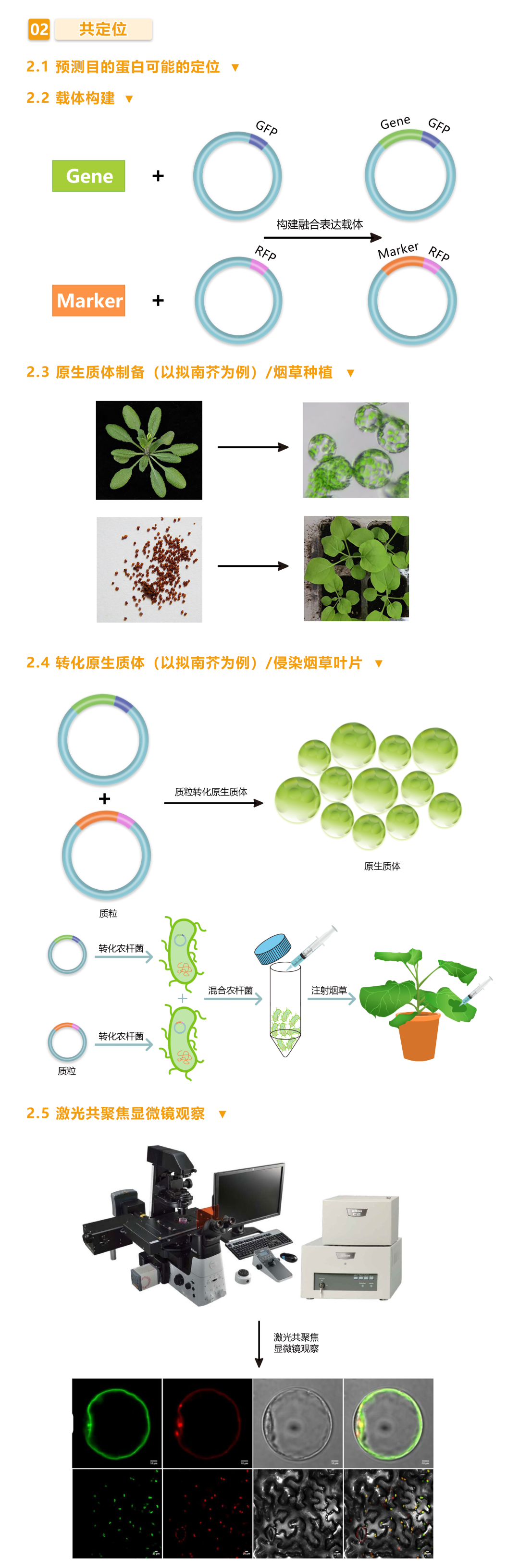
图3 伯远生物亚细胞共定位实验流程。
表1 伯远生物亚细胞定位实验中的细胞器Marker信息。
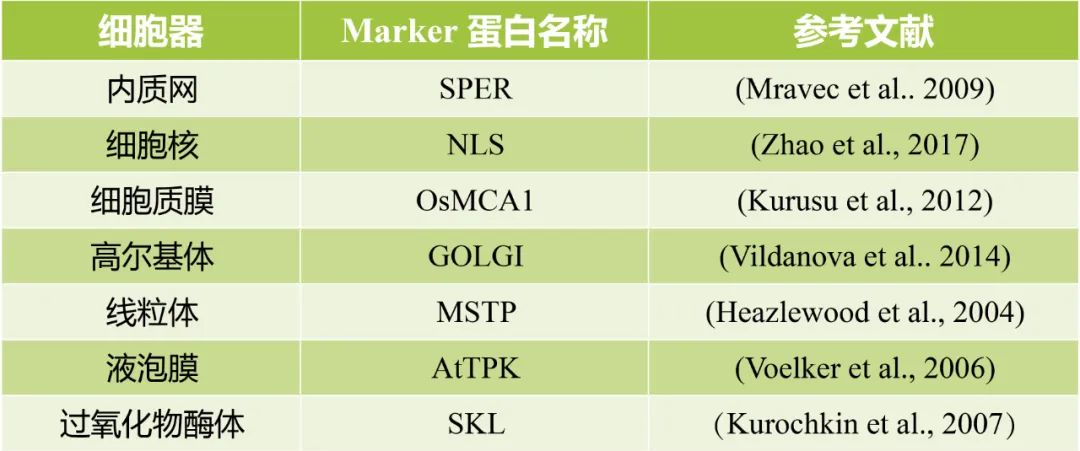
在之前的文章“再也不用担心目的基因定位在哪儿了?!!”中,小远带领大家了解了不同基因定位在不同亚细胞器的荧光图片,感兴趣的小伙伴可以去看看哟。下面给大家展示的是伯远生物的一些案例哟!
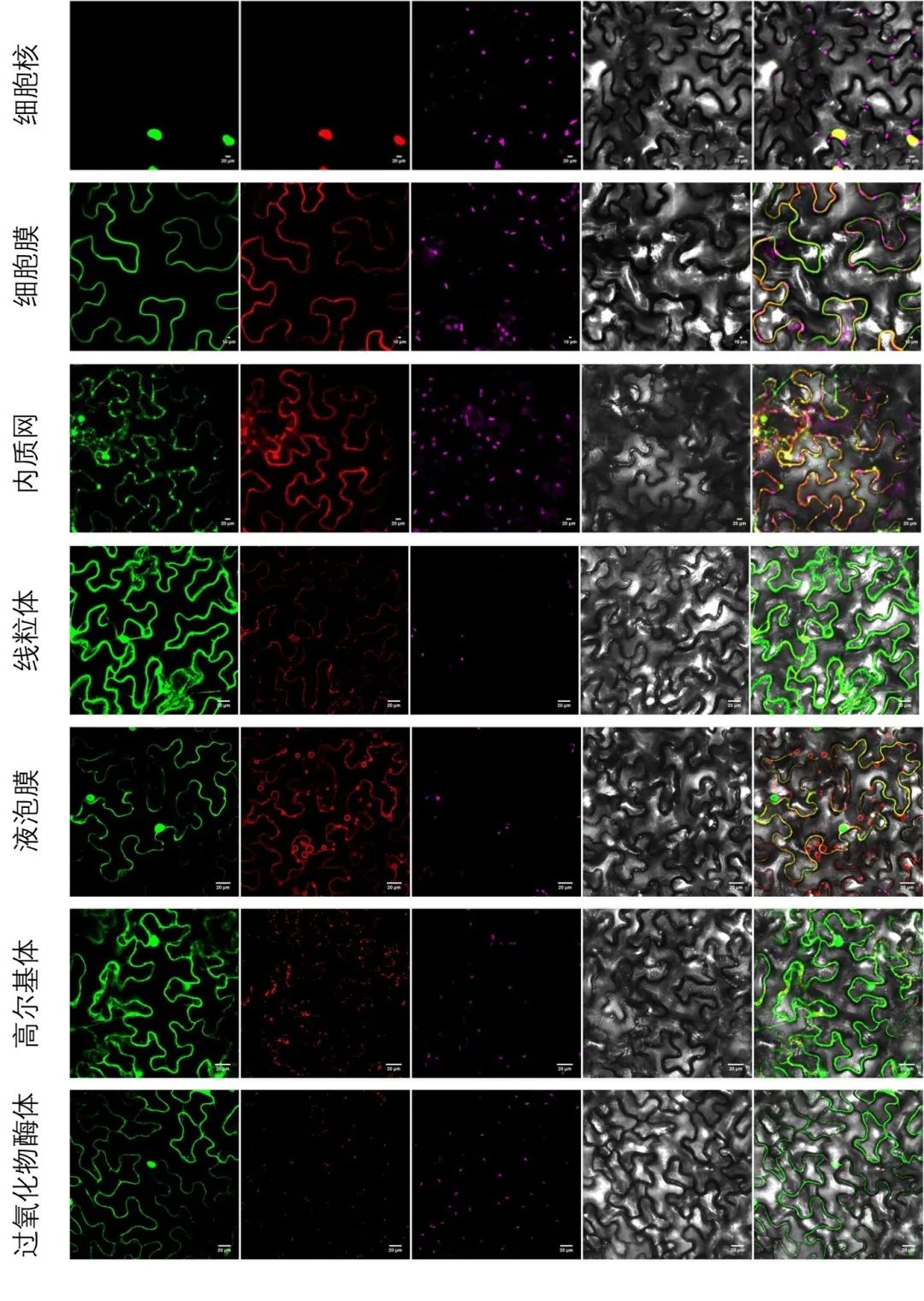
图4 以烟草叶片为材料进行亚细胞共定位。将目的蛋白基因序列构建至pBWA(V)HS-GFP载体上,将表1中相应细胞器的Marker基因序列构建至pBWA(V)HS-RFP载体上,通过农杆菌浸染法共同转化烟草叶片,观察荧光信号。
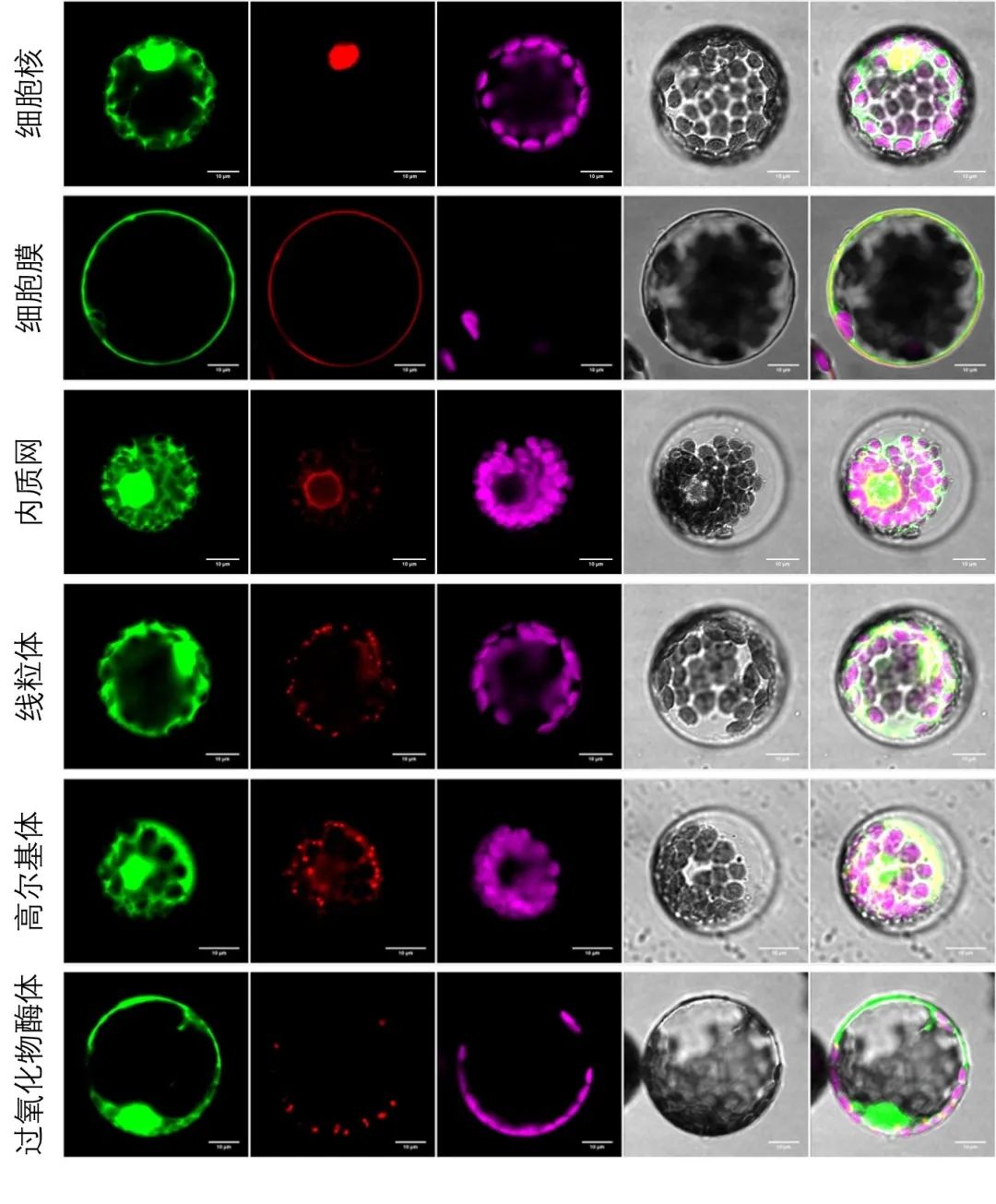
图5 以烟草原生质体为材料进行亚细胞共定位。将目的蛋白基因序列构建至pBWA(V)HS-GFP载体上,将表1中相应细胞器的Marker基因序列构建至pBWA(V)HS-RFP载体上,通过PEG介导法共同转化烟草原生质体,观察荧光信号。
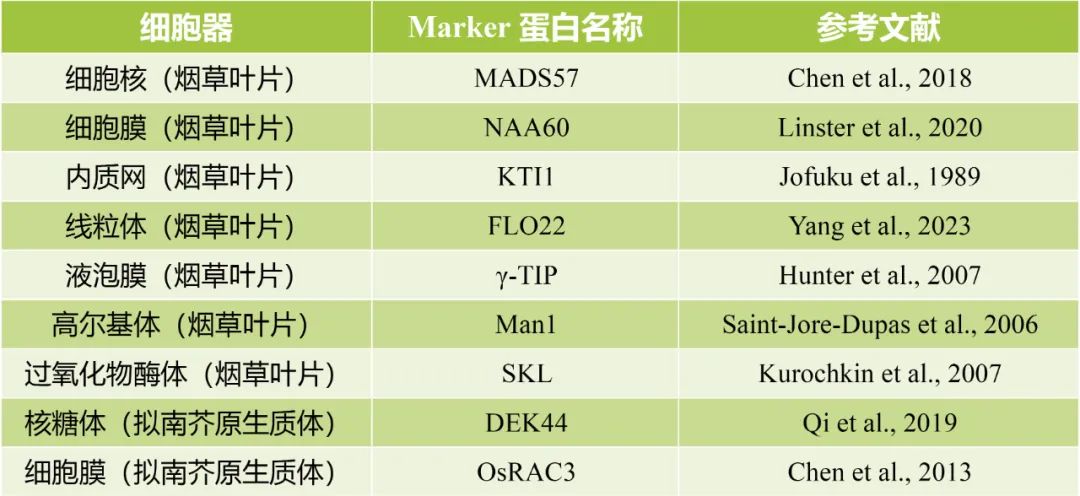
除了上表中使用的Marker蛋白外,伯远生物最近还更新了烟草叶片7个细胞器的Marker蛋白以及拟南芥原生质体2个细胞器的Marker蛋白,其相关信息如表2所示:
表2 伯远生物最近更新的
亚细胞定位实验中的细胞器Marker信息。
下面给大家展示一下将伯远生物最近更新的各种细胞器Marker质粒单独转染烟草叶片或拟南芥原生质体中观察到的结果图(图6、图7)。
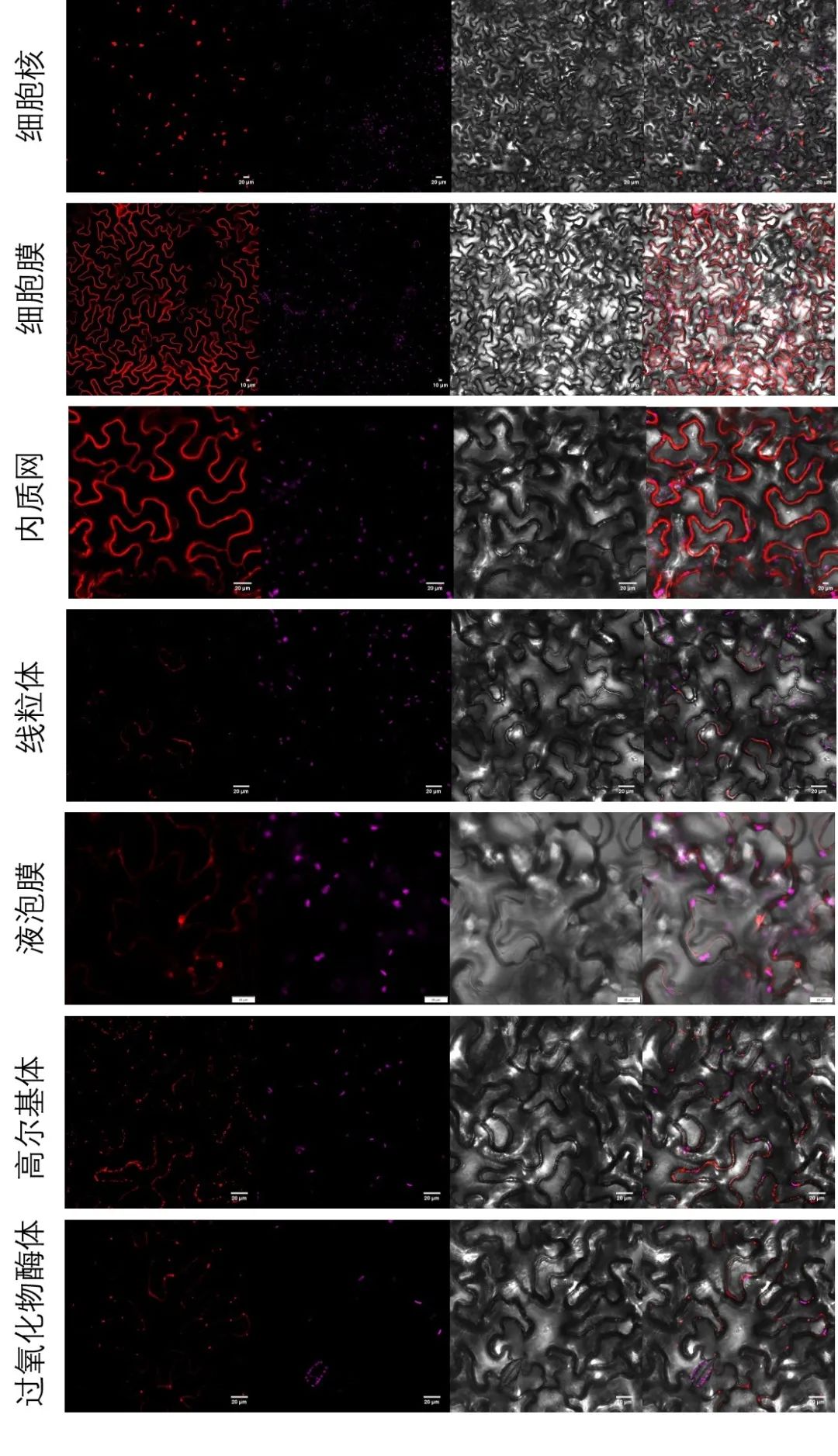
图6 表2中相应细胞器Marker载体转化烟草叶片进行荧光观察。将相应细胞器Marker基因序列构建至pBWA(V)HS-RFP载体上,通过农杆菌浸染法瞬时转化烟草叶片,观察荧光信号。
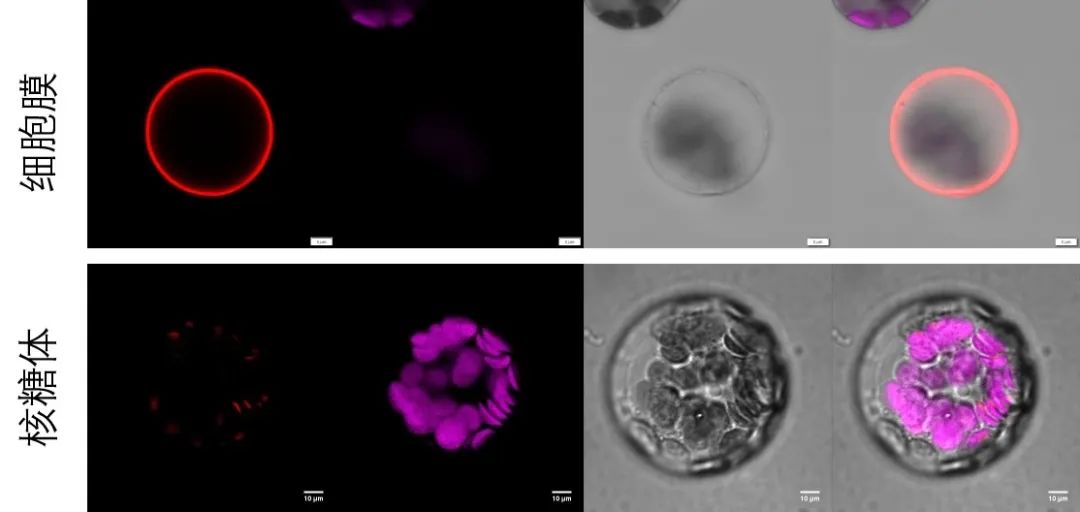
图7 表2中相应细胞器Marker载体转化拟南芥原生质体进行荧光观察。将相应细胞器Marker基因序列构建至pBWA(V)HS-RFP载体上,通过PEG介导法瞬时转化拟南芥原生质体,观察荧光信号。
除了可以在烟草叶片、烟草原生质体、拟南芥原生质体中进行亚细胞定位实验外,伯远生物还可以在水稻黄化苗和绿苗、小麦、大麦、玉米原生质体中进行亚细胞定位实验,最近还研发了大豆、紫花苜蓿原生质体的提取方法并利用其原生质体进行亚细胞定位,下面让我们一起来看看实验案例吧。
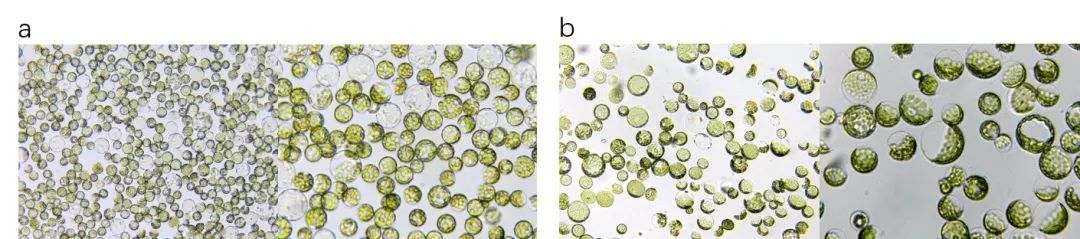
图8 大豆和紫花苜蓿原生质体提取。a.大豆原生质体;b.紫花苜蓿原生质体。
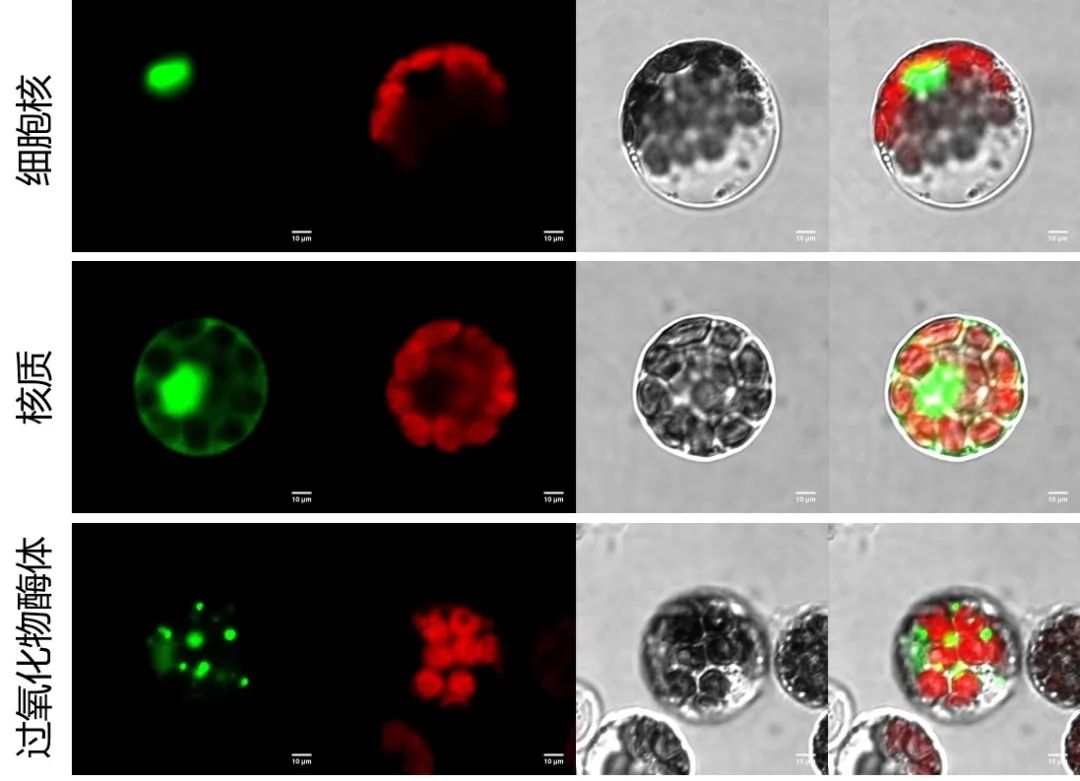
图9 以大豆原生质体为材料进行亚细胞定位。将目的蛋白基因序列构建至pBWA(V)HS-GFP载体上,通过PEG介导法瞬时转化大豆原生质体,观察荧光信号。
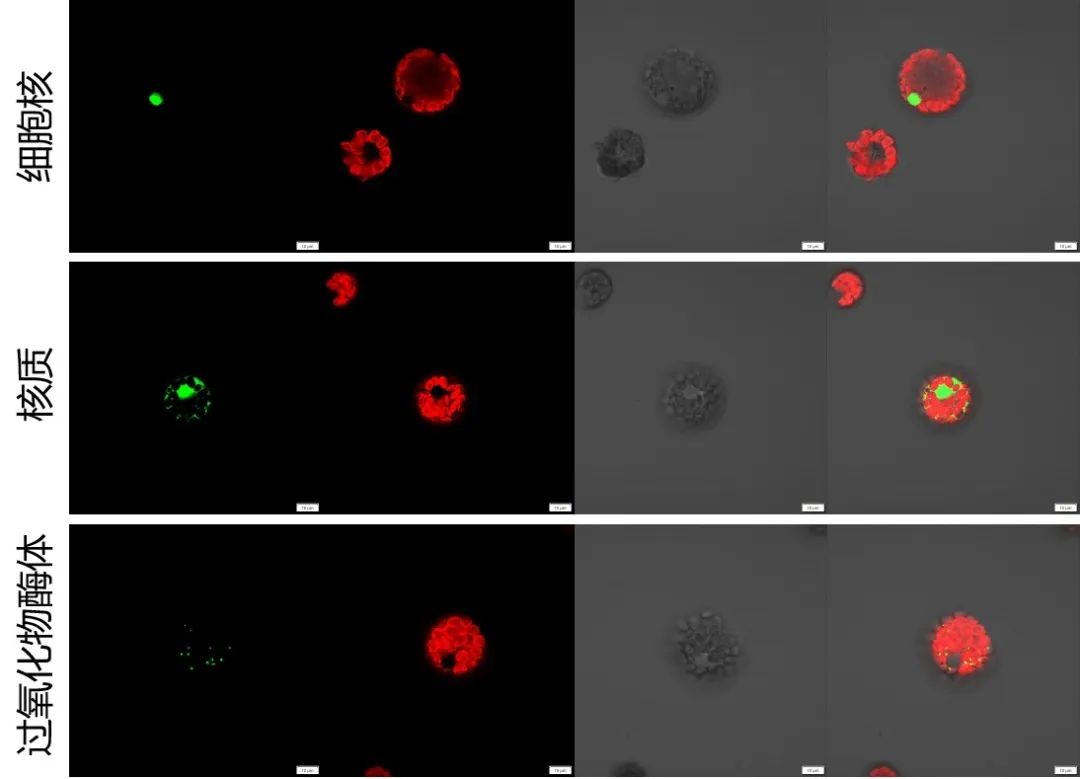
图10 以紫花苜蓿原生质体为材料进行亚细胞定位。将目的蛋白基因序列构建至pBWA(V)HS-GFP载体上,通过PEG介导法瞬时转化紫花苜蓿原生质体,观察荧光信号。
以上就是伯远生物近期对亚细胞定位实验进行的一些优化,主要包括亚细胞定位细胞器Marker的优化以及开发了新的原生质体体系(大豆与紫花苜蓿),有相关业务需求的小伙伴快来咨询吧!
表3 伯远生物亚细胞定位实验周期。

1、伯远生物采用已被授权的国际PCT专利技术(专利号:US 10,144,936 B2)进行亚细胞定位载体的构建,且该组装为无缝组装,可排除非必要的序列干扰实验结果。
2、伯远生物建立了丰富的亚细胞定位载体资源,包含了基于红色荧光蛋白、黄色荧光蛋白、绿色荧光蛋白、青色荧光蛋白的载体库,方便选择使用。
3、拥有细胞核、细胞膜、液泡膜、内质网、过氧化物酶体、高尔基体、线粒体多种细胞器的Marker载体(mCherry荧光),以方便共定位时选择。
4、可瞬转烟草叶片和多种原生质体(水稻黄化苗和绿苗、拟南芥、小麦、大麦、烟草、玉米、大豆、紫花苜蓿)进行亚细胞定位观察。
5、常备水稻、烟草、小麦、玉米、拟南芥等的原生质体以及烟草叶片,每周定期观察,可快速获得观察结果。
6、每个基因拍5-6个细胞,保证实验的可靠性与可重复性。
构建亚细胞定位载体时,GFP融合位置为什么有N端、C端之分?
为什么不同的受体材料有时得到的定位结果不一样?
用水稻原生质体进行亚细胞定位时,为什么不拍摄叶绿体的自发荧光?
为什么要提供GFP空载的对照图片?
为什么有的共定位图片中叶绿体通道是玫红色?
为什么有的基因定位结果与预想的不一致?
为什么有的普通定位结果出来后无法准确判断其定位位置?
为什么不先对一个基因做预测,在预测的结果上直接做共定位?
为什么有的基因做原生质体转化时荧光蛋白不亮?
拍摄时为什么有明场通道?
以烟草叶片为材料进行亚细胞定位时,液泡膜、细胞膜、细胞质三者怎样区分?
为什么在构建载体时直接默认选择大载体,不告诉可以用小载体?
烟草叶片瞬时转化效率低,怎么解决?
瞬时转化原生质体进行亚细胞定位实验需要注意什么?

