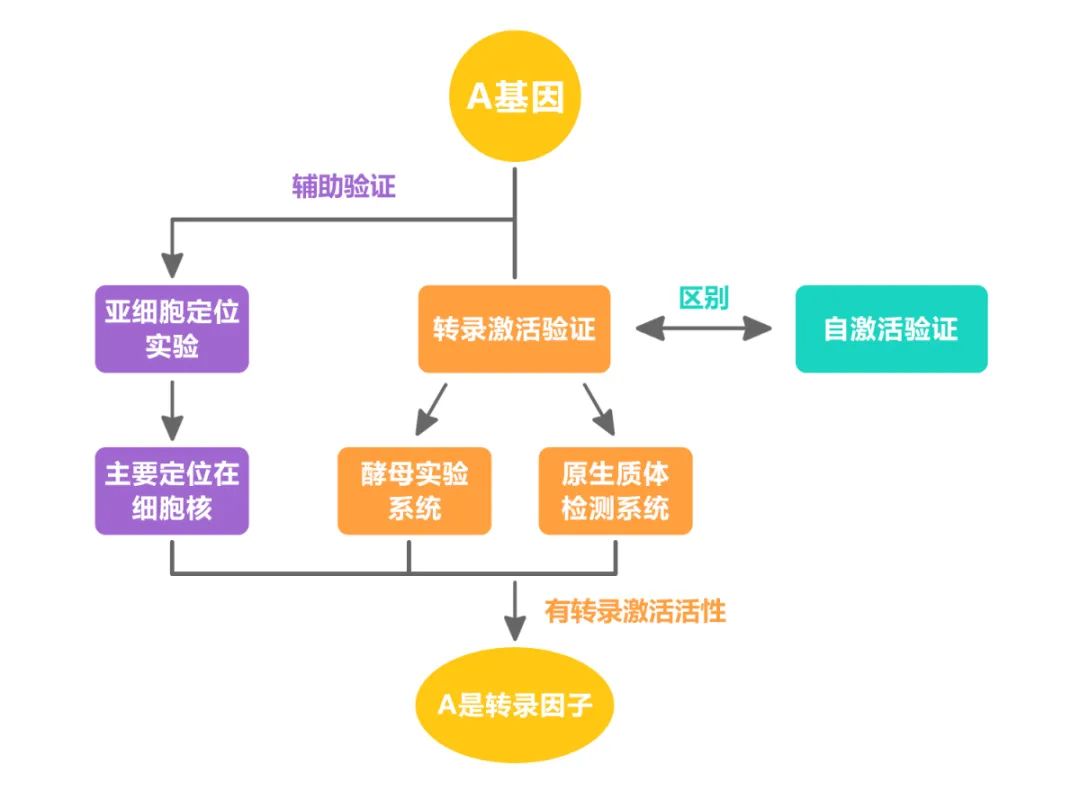
还记得有人问过伯小远这样一个问题:如何验证一个基因是转录因子?为了系统地回答这个问题,伯小远去找了一些资料,整理了下面的文章,希望大家看完之后在文末踊跃留言,发表一下自己的看法哟,伯小远真的好想和大家互动呀!
假如你得到了一个基因,并且猜测该基因可能是转录因子,那么通过什么实验来进行验证呢?其实要验证一个基因是转录因子,可以从两个方面入手,其一是亚细胞定位分析,其二是转录激活分析,下面我们就通过具体的文献来给大家展示一下相关的方法。
首先,我们先来回顾一下转录因子的定义:在分子生物学中,转录因子(Transcription factor)是指能够结合在某基因上游特异核苷酸序列上的蛋白质,这些蛋白质可通过调控核糖核酸聚合酶(RNA聚合酶)与DNA模板的结合,参与基因的转录过程,即转录因子的本质是与DNA特异性结合的一系列蛋白质。根据定义,不难看出转录因子调控基因转录的过程发生在细胞核中,因此通过研究其亚细胞定位情况,在一定程度上就可以证明其是否为转录因子。
在“Drought-responsive WRKY transcription factor genes TaWRKY1 and TaWRKY33 from wheat confer drought and/or heat resistance in Arabidopsis”一文中,TaWRKY1和TaWRKY33作为干旱WRKY转录因子,它们的亚细胞定位结果都定位在细胞核中。
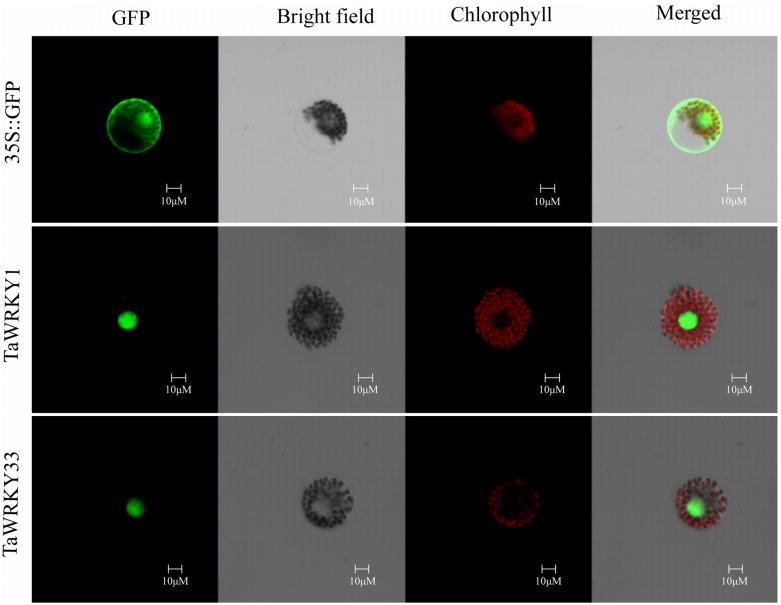
图1 TaWRKY1和TaWRKY33蛋白的亚细胞定位(He et al., 2016)。
转录因子定位在细胞核中,这个结果符合我们对转录因子的一般认识。但是如果你研究的一个转录因子,它的亚细胞定位不在细胞核中或不完全定位在细胞核中,这个时候你也不要惊讶,请看以下两个栗子。
在“Mobility and localization of the iron deficiency-induced transcription factor bHLH039 change in the presence of FIT”一文中,在烟草表皮细胞中观察蛋白的亚细胞定位,发现单独表达FIT-GFP时,FIT在烟草表皮细胞中具有细胞核和细胞质的双重定位,只不过细胞质中的信号比较弱(图2a);单独表达bHLH039-mCherry时,转录因子bHLH039主要定位在细胞外周,细胞核中有微弱的信号(图2b)(原文中是这样描述的,其实伯小远真的没发现在细胞核中有信号,不过这里还是尊重作者的意思);当bHLH039-mCherry与FIT-GFP共表达时,bHLH039-mCherry向核定位方向发生了显著地转移,但FIT-GFP的定位并不会受bHLH039-mCherry的影响(图2c);当bHLH039-mCherry与GFP共表达时,单独GFP的存在并不会影响bHLH039-mCherry的定位(图2d)。
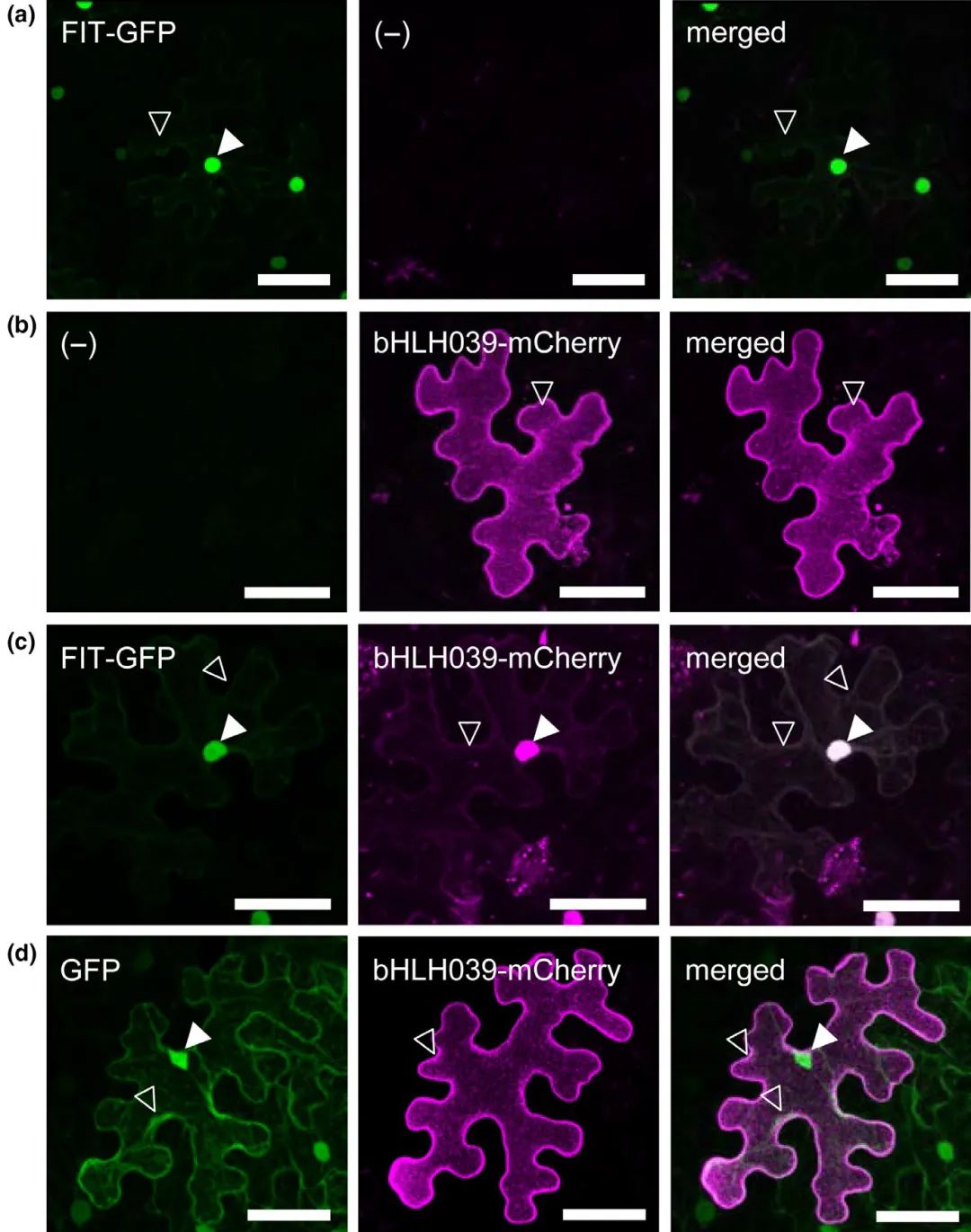
图2 bHLH039的亚细胞定位依赖于烟草叶片表皮细胞中FIT的存在(Trofimov et al., 2019)。
在野生型拟南芥和缺失FIT的突变体拟南芥的原生质体中观察亚细胞定位,发现在野生型拟南芥的原生质体中单独表达bHLH039-mCheery时,其亚细胞定位结果显示细胞核和细胞质的双重定位(图3a),而共表达FIT-GFP与bHLH039-mCheery时,bHLH039-mCheery显著向细胞核转移(图3b),结果与烟草表皮细胞中的一致;在缺失FIT的突变体拟南芥原生质体中,单独表达bHLH039-mCheery时,其荧光信号在细胞外周(图3c),而共表达FIT-GFP与bHLH039-mCheery后,恢复了bHLH039的核积累(图3d)。
以上,通过烟草叶片表皮细胞和拟南芥原生质体系统证明了FIT对bHLH039核积累的重要性。
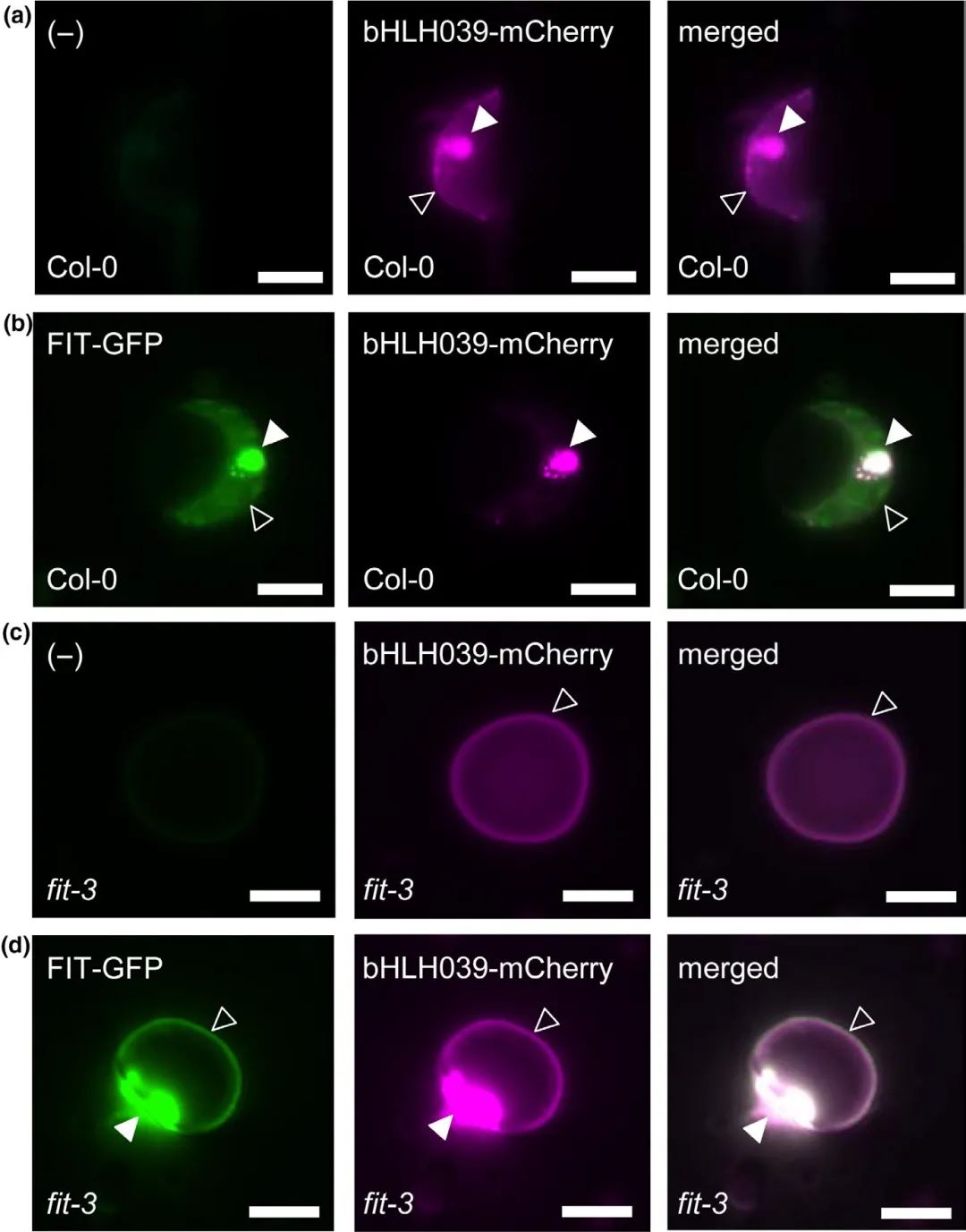
图3 拟南芥原生质体中bHLH039依赖FIT的亚细胞定位(Trofimov et al., 2019)(Col-0表示野生型拟南芥,fit-3表示缺失FIT的拟南芥突变体)。
通过这个栗子可以看出, bHLH039作为一个转录因子,单独转化烟草细胞或拟南芥原生质体时,其亚细胞定位都没有显示完全的细胞核定位(图2b和图3a),因此对于转录因子定位在细胞核中并不是一个绝对的结论,还是存在很多例外的!
在这个栗子中,我们还可以看出,用不同的受体材料做亚细胞定位分析,其结果可能存在不一致的情况,比如bHLH039在烟草叶片细胞中主要定位在细胞质中(图2b),而在拟南芥原生质体中细胞核和细胞质都有比较明显的信号(图3a),所以,大家以后不要再问伯小远用不同的材料做亚细胞定位结果为什么会不一致这个问题了,看这里,你就会有答案了!
最近发表在PANS上的“Nucleocytoplasmic trafficking and turnover mechanisms of BRASSINAZOLE RESISTANT1 in Arabidopsis thaliana”一文解析了BR调节BZR1核质穿梭和蛋白稳定性的分子机理,进一步完善了BR信号转导调控的分子机制,内容有点复杂,先介绍一下背景,再来聊我们想要说明的!
油菜素甾醇(Brassinosteroids,BRs)是一种重要的植物激素,参与调节植物的生长、发育以及胁迫响应等多个生物学过程。经过多年的研究,BR信号转导分子机制得到了较清楚的解析:BRs可被质膜定位受体BRI1及其共受体BAK1感知,BR激活的BRI1可通过连续磷酸化和去磷酸化失活,促进细胞蛋白激酶BIN2的降解。当BR信号通路失活时,BIN2磷酸化BZR1家族转录因子(BZRs),从而阻止它们调节其靶基因的表达,进一步在14-3-3蛋白的帮助下,BIN2的磷酸化促进了BZR1的核输出,从而导致磷酸化的BZR1在细胞质中的积累;当BR信号通路激活时,BIN2被灭活和降解,进而通过蛋白磷酸酶PP2A促进BZR1的去磷酸化,致使去磷酸化的BZR1在细胞核中积累,在那里它与下游BR调控基因的启动子结合,调节它们的转录。因此,BZR1的磷酸化和去磷酸化,以及核质定位的调节是BR信号转导的核心事件。这里我们就单独聊一聊BR如何影响BZR1的亚细胞定位。这里作者用到了一种比较好的方法,就给大家介绍仔细一点,学习一下高分文章都是怎么做实验的。
虽然BR处理促进了BZR1的核积累,但不清楚BR处理后,在细胞核中积累的BZR1是重新合成的还是细胞质原有的,或者两者都有。要确定这一点,需要有方法区分新合成的BZR1(在BR处理后)和已经存在的BZR1(在BR处理前),并分别跟踪细胞内的每个BZR1。
为了达到这个目的,作者使用了Kaede,一种从石质珊(Trachyphyllia geoffroyi)中分离出来的光转换荧光蛋白,该蛋白在正常条件下可在488nm的激发光下发出绿色荧光(Kaedegreen);而紫外光(390-405nm)的照射会导致Kaede蛋白发生不可逆的裂解,裂解后的Kaede蛋白的激发光转变为543nm,发射光由绿色转变为红色(Kaedered)。因此利用Kaede标签可以有效区分紫外光处理后新合成的BZR1-Kaedegreen蛋白(绿色)和原有的BZR1-Kaedered蛋白(红色)。
作者利用含有BZR1pro:BZR1-Kaede-FLAG转基因植株(基因自己的启动子驱动自己的基因组序列融合标签的载体)进行实验。共聚焦显微镜和免疫印迹法显示,与野生型BZR1-YFP一样,BZR1-Kaedegreen融合蛋白在没有BR的情况下被磷酸化并定位于细胞质,但在油菜素内酯(BL)处理30min后其被去磷酸化并定位于细胞核(图4和图5A)。幼苗在543nm的激发光下,只观察到背景荧光(图5B);而在405nm紫外光照射幼苗5min后,BZR1-Kaedegreen荧光信号急剧下降(图5C),并且细胞质出现较强的BZR1-Kaedered荧光信号(图5D),这表明5min的紫外光照射并没有改变BZR1-Kaede的亚细胞定位。因此,作者接下来用1μM eBL处理经紫外光照射的幼苗30min,观察到绿色(残留的未转化和新合成的BZR1-Kaedegreen蛋白的混合物)和红色(转化的BZR1-Kaedered蛋白仅在BR处理前)荧光信号的强烈积累(图5E和F)。以上结果表明,胞质BZR1在BR通路激活后,细胞质中原有的BZR1可以被招募到细胞核中。
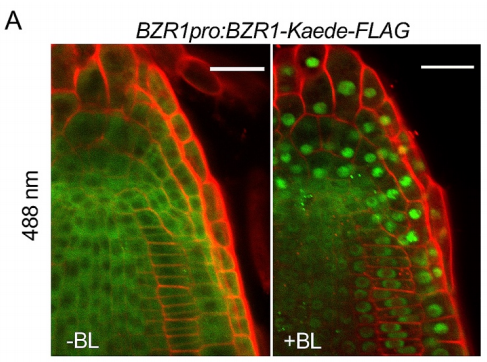
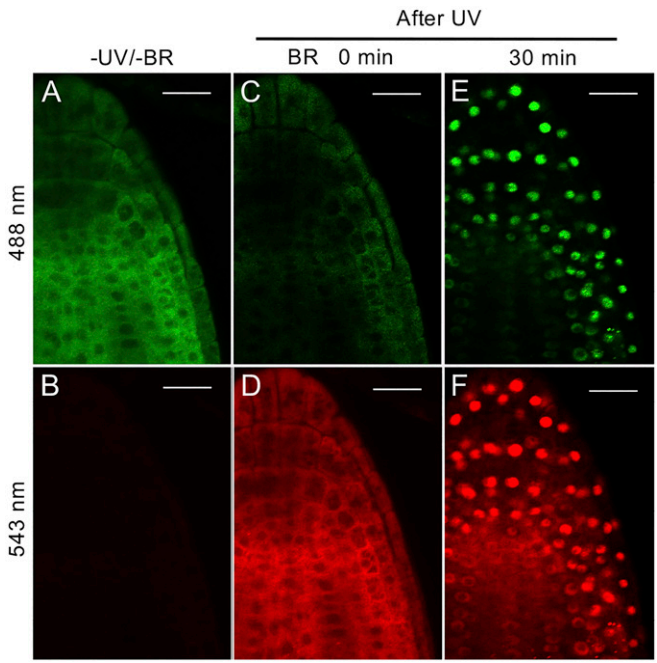
图5 BR将胞质BZR1招募到细胞核(Wang et al., 2021)。
BZR1作为转录因子,在没有经过任何处理的时候,其亚细胞定位也是定位在细胞质中的,经过BL处理之后会被招募到细胞核中,这与栗子一有异曲同工之妙,只不过这个实验研究的会更深入一些,解释了细胞核中的成分是新合成的还是原本细胞质中的成分进行了转移这一问题,结果表明是细胞质中原有的成分进行了转移。这个方法大家可以学起来哦!
好了,关于转录因子亚细胞定位的实验就介绍这么多啦,大家只要记住转录因子的亚细胞定位一般是定位在细胞核中的,但是也有例外,上面的两个栗子就是很好的说明。
下面我们来聊一聊转录因子的转录激活分析,在讲这个之前,我们先来回顾一下酵母双杂的定义,这对于大家理解转录因子的转录激活分析会有帮助哦!
酵母双杂是基于对真核细胞转录因子特别是酵母转录因子GAL4性质的研究。GAL4包括两个彼此分离但功能必需的结构域。位于N端1-174位氨基酸残基区段的DNA结合域(DNA binding domain, DNA-BD)和位于C端768-881位氨基酸残基区段的转录激活域(Activation domain, AD)。DNA-BD能够识别位于GAL4效应基因(GAL4-responsive gene)的上游激活序列(Upstream activating sequence, UAS),并与之结合。而AD则是通过与转录机构(Transcription Machinery)中的其他成分之间的结合作用,以启动UAS下游的基因进行转录。DNA-BD和AD单独作用时不能激活转录反应,但是当二者在空间上充分接近时,则呈现完整的GAL4转录因子活性并可激活UAS下游启动子,使启动子下游基因得到转录。
对于转录因子的转录激活分析,就是将需要验证的转录因子构建到BD载体上,不需要构建AD载体,如果转录因子具有转录激活活性,那么下游的报告基因就能被激活表达,这样也就验证了转录因子是具有转录激活活性的。另外,在酵母双杂实验中,自激活验证用到的也是这个原理,为了避免要验证的两个蛋白之间的互作存在假阳性,在进行双杂实验之前会对构建好的BD载体进行自激活分析,就是避免BD载体上连接的是转录因子,从而导致假阳性的结果。下面我们先来看一看转录因子的转录激活分析,这里介绍两种验证转录因子转录激活活性的方法。
在“Plant NAC-type transcription factor proteins contain a NARD domain for repression of transcriptional activation”一文中,作者用到了多种方法对转录因子的转录激活活性进行了验证,下面我们就去学习一下!
作者利用酵母实验系统分析了大豆NAC家族3个基因GmNAC11、GmNAC20和GmNAC35的转录激活能力。首先,将3个基因的编码区分别融合到GAL4的DNA结合区域,生成pBD-GmNAC融合质粒;然后,将质粒转化到酵母株YRG-2中,并检测其在GAL4上游激活序列激活转录的能力,以及在无组氨酸培养基中促进酵母生长的能力,以含有pGAL4和pBD载体的转化子分别作为阳性和阴性对照。如图7所示,所有的转化子在正常培养基(YPAD)上都能良好生长;在不含组氨酸的SD培养基上,含有GmNAC11、GmNAC35和pGAL4的转化子生长良好,而含有GmNAC20和pBD的酵母则不能在同一培养基上生长,并且GmNAC11、GmNAC35或pGAL4细胞中β-半乳糖苷酶活性的X-gal染色呈蓝色(图7)。这些结果表明,GmNAC11和GmNAC35具有转录激活能力,而GmNAC20不具有转录激活能力。
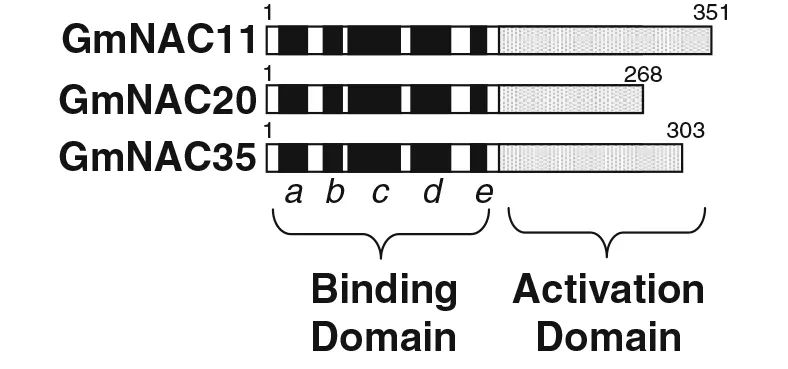
图6 三个GmNAC基因的结构示意图(Hao et al., 2010)。
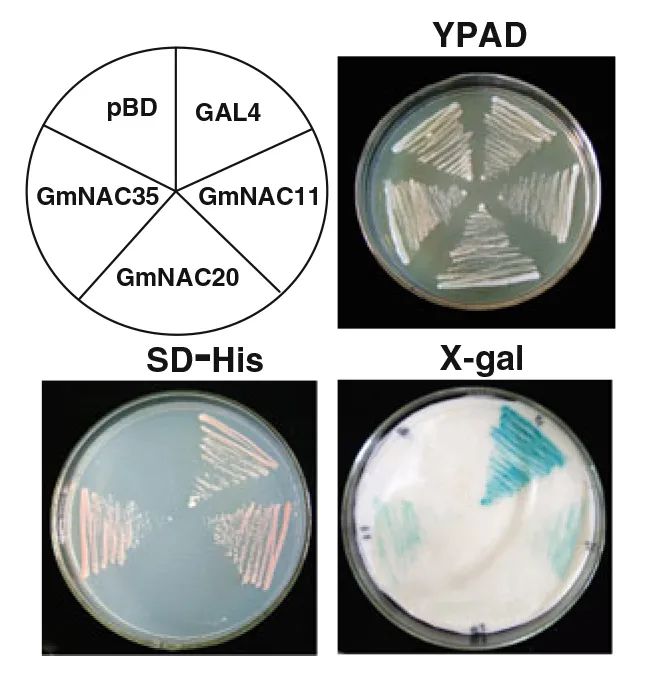
图7 GmNAC蛋白的转录激活活性(Hao et al., 2010)。
对于转录因子GmNAC20没有转录激活能力,作者进行了探究,分别构建了包含N端NAC DNA结合域(氨基酸编号:1–175, 1/175)和C端推测的激活域(氨基酸编号:162–268, 162/268)的截短载体,并通过观察SD-His的生长状态和β-半乳糖苷酶的活性来探索它们的转录激活能力,发现含有GmNAC20 C端区(162/268)的酵母在选择培养基上生长良好,呈阳性蓝色,β-半乳糖苷酶活性强,说明该区域具有较强的转录激活能力;相反,NAC的N端结构域(1/175)没有任何转录激活活性(图8)。
由于GmNAC20的C端区(162/268)具有转录激活能力,而GmNAC20的全长不具备转录激活能力,因此作者提出,在全长蛋白中可能存在一个抑制域来干扰C端激活域的作用。通过将N端结构域的各个部分拆分与C端进行组合验证,最终找到GmNAC20 C端激活域的抑制区域位于100-134氨基酸区域内(图8)。

图8 GmNAC20转录激活活性的截断分析(Hao et al., 2010)。
将GmNAC20的抑制结构域记为NARD,作者想要验证该抑制结构域是否也会抑制其他家族的转录因子,因此选择了不同家族的4个转录因子Dof、WRKY、AP2/DRE和NAC进行检测。作者在这里用到了两种方法,第一种方法是将NARD融合到4个蛋白的N端,利用酵母系统进行验证,上面的实验中介绍过该方法,这里就不再介绍。下面介绍第二种方法——原生质体检测系统。以VP16转录激活域作为阳性对照,GAL4DBD作为阴性对照,以NARD与VP16的N端或C端融合的融合蛋白作为效应因子,检测NARD的抑制功能(图9a)。以TATA盒前5个GAL4结合元件的35S启动子驱动的Firefly荧光素酶基因为报告基因,以35S启动子驱动的Renilla荧光素酶基因为内参基因(图9a),即Effectors中的5个载体分别与Reporter的载体共转化原生质体。结果显示VP16相对荧光素酶活性高于对照GAL4DBD,而在VP16的N端或C端添加NARD可以抑制80%以上的VP16活性,含有两个VP16拷贝的效应子载体作为融合蛋白分子内两种蛋白相互作用的阳性对照(图9b)。这些结果表明,在拟南芥原生质体检测系统中,NARD可以作为一个抑制域,其结果与酵母系统的一致。
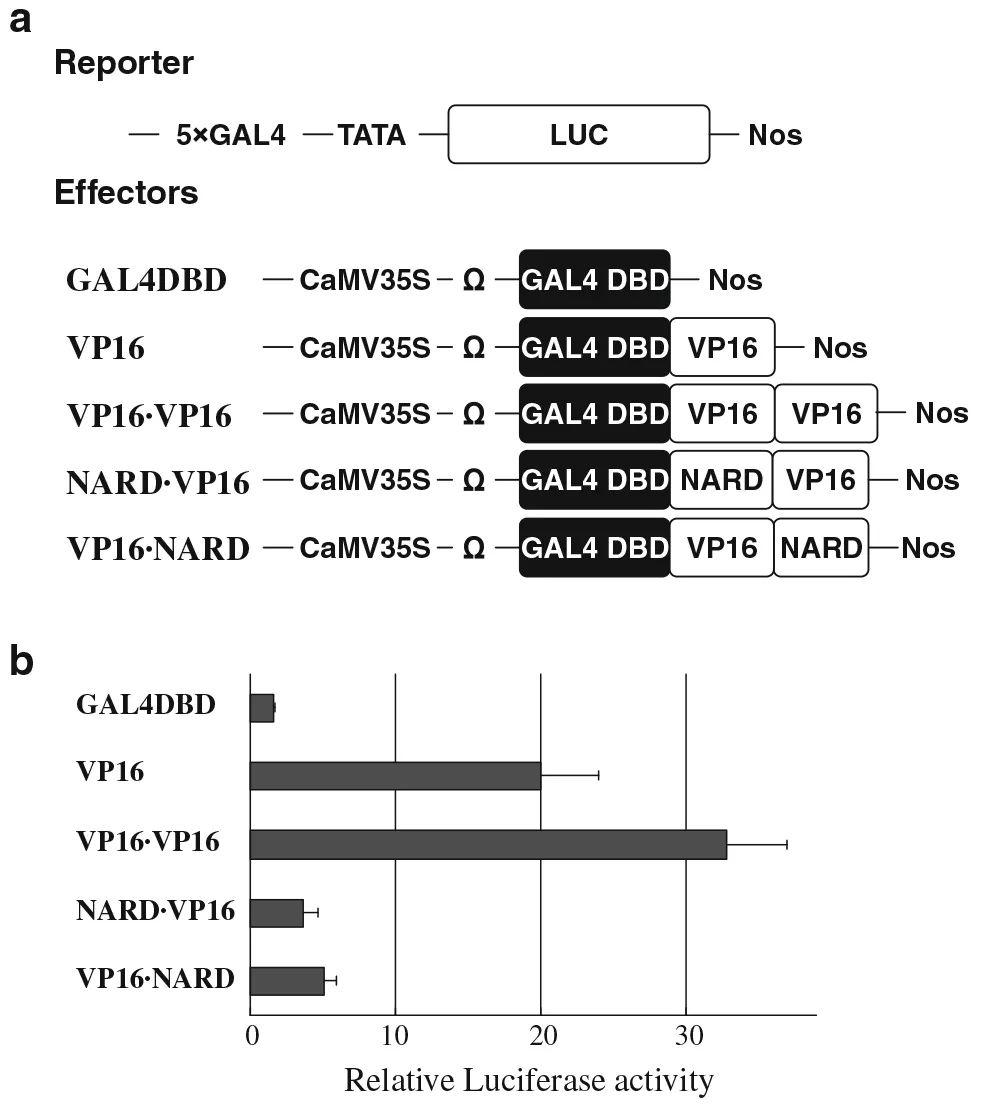
图9原生质体实验中NARD对VP16活性的影响(Hao et al., 2010)。
酵母实验系统与原生质体检测系统都可以检测转录因子的转录激活活性,只不过前者是以β-半乳糖苷酶的活性来衡量,后者是以荧光素酶活性来衡量,具体使用什么方法自己选择;
全长转录因子没有转录激活活性,其原因可能是全长转录因子内部存在转录抑制结构域,可以去探究其具体的位置。
自激活实验
最后简单讲一下酵母双杂实验中的自激活实验,一般BD可单独与GAL4上游激活序列UAS结合,但不能引起转录。如果将一段具有转录激活活性的转录因子构建到BD载体上,若其表达产生的融合BD单独与UAS结合后能引起下游报告基因的转录,则说明诱饵基因具有自激活现象。
在进行酵母双杂实验之前,首先需要将连在BD载体上的bait基因进行自激活验证,如果验证之后不存在自激活现象,就可以进行后续的实验。如下图所示,3组实验在二缺板(DDO)上都能正常生长,但是在三缺板(TDO,添加了X-α-gal)上只有阳性对照可以正常生长,阴性对照和自激活验证组都不能正常生长,说明bait基因不具有自激活活性,可以进行后面的双杂实验。

图10 酵母双杂实验自激活验证(图片来源:伯远生物)。
文章到这里就结束啦,对于如何验证一个基因是转录因子,伯小远已经找了一些具有代表性的文献给大家进行了说明,以及对于一些名字相似的概念也进行了区分,希望大家看完可以收获满满哟!
思路小结
Hao Y J, Song Q X, Chen H W, et al. Plant NAC-type transcription factor proteins contain a NARD domain for repression of transcriptional activation[J]. Planta, 2010, 232(5): 1033-1043.
He G H, Xu J Y, Wang Y X, et al. Drought-responsive WRKY transcription factor genes TaWRKY1 and TaWRKY33 from wheat confer drought and/or heat resistance in Arabidopsis[J]. BMC plant biology, 2016, 16(1): 1-16.
Trofimov K, Ivanov R, Eutebach M, et al. Mobility and localization of the iron deficiency‐induced transcription factor bHLH039 change in the presence of FIT[J]. Plant Direct, 2019, 3(12): e00190.
Wang R, Wang R, Liu M, et al. Nucleocytoplasmic trafficking and turnover mechanisms of BRASSINAZOLE RESISTANT1 in Arabidopsis thaliana[J]. Proceedings of the National Academy of Sciences, 2021, 118(33).

