单倍体育种(Haploid breeding)是植物育种手段之一。通过单倍体诱导获得的单倍体植株经染色体加倍后即可出现纯系植株(2n),从中选出的优良纯系后代不分离,植株表型整齐一致,并且与传统育种(6-8代自交)相比可显著缩短育种年限,因此具有重要的意义。该技术不仅适合某些特定表型的纯合,例如,自交不亲和植物的育种,也可以应用于快速繁殖、突变育种和转基因育种等方面。随着单倍体技术的不断革新,目前该技术已经成为现代育种技术体系的三大核心技术之一。今天我们主要是以玉米单倍体育种为主线,结合单倍体育种在其他物种中的拓展应用来和大家一起探讨,单倍体育种的技术流程、最新进展以及应用现状,接下来,让我们一起进入今天的主题吧~
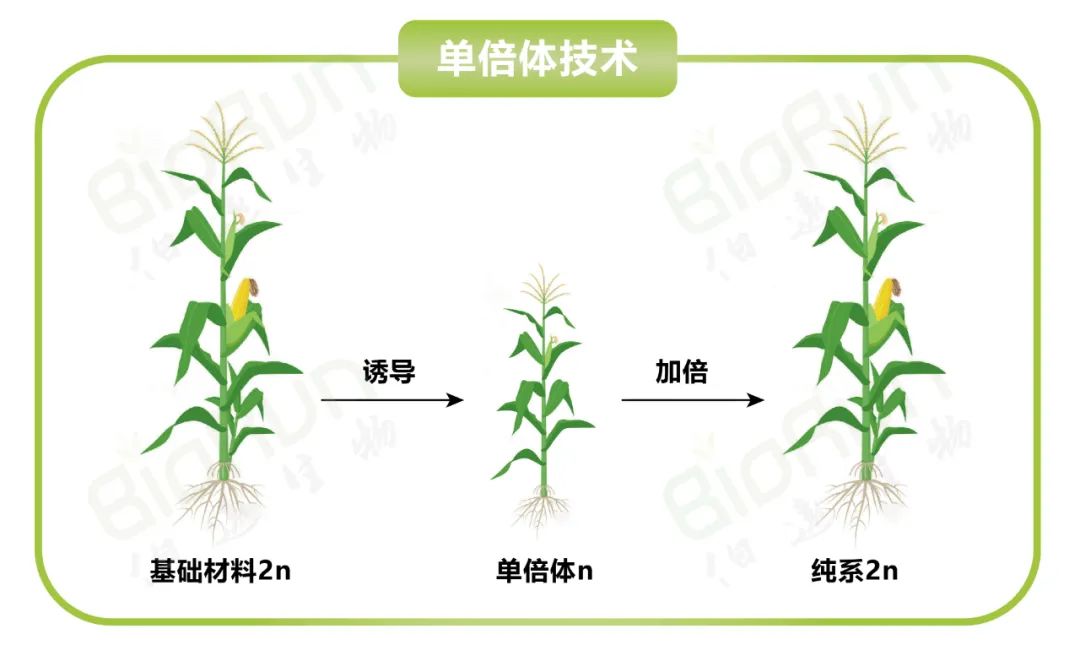
图1 单倍体育种(图片来源:伯远生物科研绘图团队)。
单倍体育种的核心是为了充分的利用杂种优势,而利用杂种优势的重要一环就是获得纯系来作为杂交品系的亲本。因此单倍体的获得是该技术能够顺利应用的核心环节。除自然产生的极少部分之外,单倍体主要是通过人工诱导产生的。人工产生单倍体的两大途径,分别是体外单倍体的诱导和体内单倍体的诱导。体外诱导方法包括花药/小孢子、胚珠/子房的离体培养,但这种方法成本高,劳动强度大,对基因型的依赖程度高,且组培过程中发生突变的概率高,因此应用范围较狭窄。体内诱导方法包括单性生殖、种间杂交和种内杂交这几类。其中以种子为基础的种内杂交方法具有劳动强度低,成本相对较低的特点,已逐渐成为玉米等物种育种的关键性技术!因此,在接下来的单倍体育种流程中,小远会重点为大家讲解以种内杂交为基础的单倍体育种。
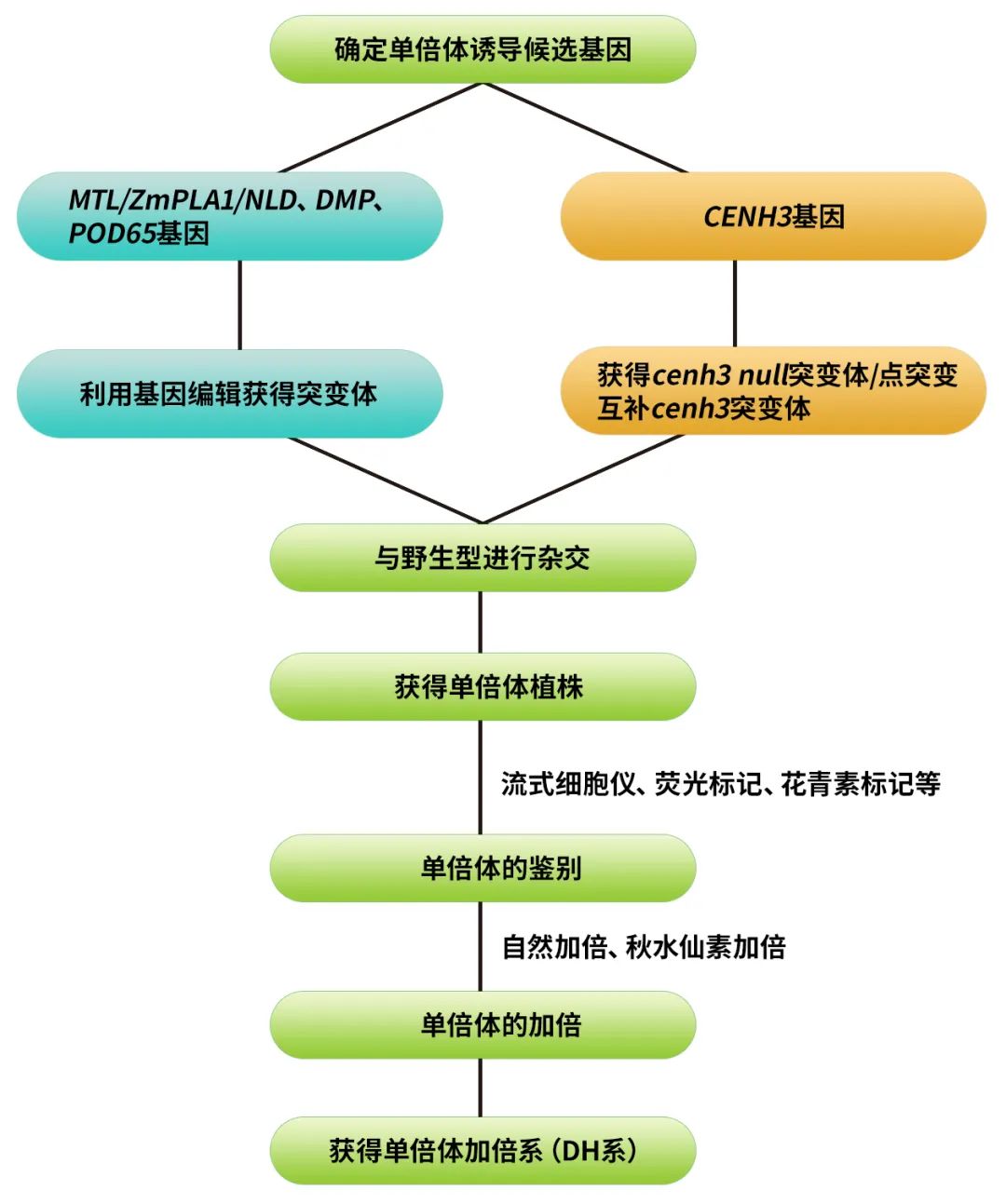
单倍体育种的基本流程可以简单概括为:单倍体诱导、单倍体鉴别、单倍体加倍这三部分。
以种内杂交为基础的单倍体诱导主要可通过:(1)单倍体诱导系诱导或(2)CENH3介导的单倍体诱导来获得相应的单倍体。
其中,单倍体诱导系的发现可以追溯到1959年,当时密苏里大学的教授Edward H. Coe发现他的一个玉米品系Stock6在作为父本与不同的母本杂交时可产生2-3%的单倍体,因此Edward将Stock6命名为“单倍体诱导系”(Coe, 1959)。在接下来的几十年中,由Stock6衍生的一系列诱导系逐渐成为获得单倍体最有效的方法。近年来,随着科学家们对Stock6诱导产生单倍体的生物学机制不断研究(图2),在2017年第一个单倍体诱导关键基因ZmPLA1/MTL/NLD被全球多家单位先后克隆,成功地揭示了单倍体诱导现象的谜底(Kelliher et al., 2017; Liu et al., 2017; Gilles et al., 2017)。2019年陈绍江团队在非Stock6材料中克隆了玉米单倍体诱导的另一个关键基因ZmDMP,并揭示了现代单倍体诱导系由原本2%的诱导率能够提升到约10%的问题(Zhong et al., 2019)。基于这些基因的功能保守性,在单子叶植物如:小麦、水稻、谷子中通过敲除ZmPLA1/MTL/NLD的同源基因,可以实现这些作物的单倍体诱导;在双子叶植物如:拟南芥、蒺藜苜蓿、番茄、烟草等中通过敲除ZmDMP的同源基因,也建立起了这些物种的单倍体诱导体系。有关这一部分的拓展研究大家想要了解的更详细的话可以查阅小远的往期文章:纯系养成——单倍体诱导技术(一)。
科学家们对单倍体诱导系的研究并没有因为取得上述成果而放慢脚步。近期,严建兵团队和李响团队通过深入解析玉米单倍体诱导关键基因ZmPLA1导致单倍体发生的分子机制,成功鉴定了一个诱导单倍体发生的新基因ZmPOD65,将该基因进行编辑后,在其后代中发现了高至7.7%的单倍体诱导率,并且该基因在双单子叶植物间的保守性,为其它作物上发展单倍体诱导技术提供了新的基因资源(Jiang et al., 2022)。
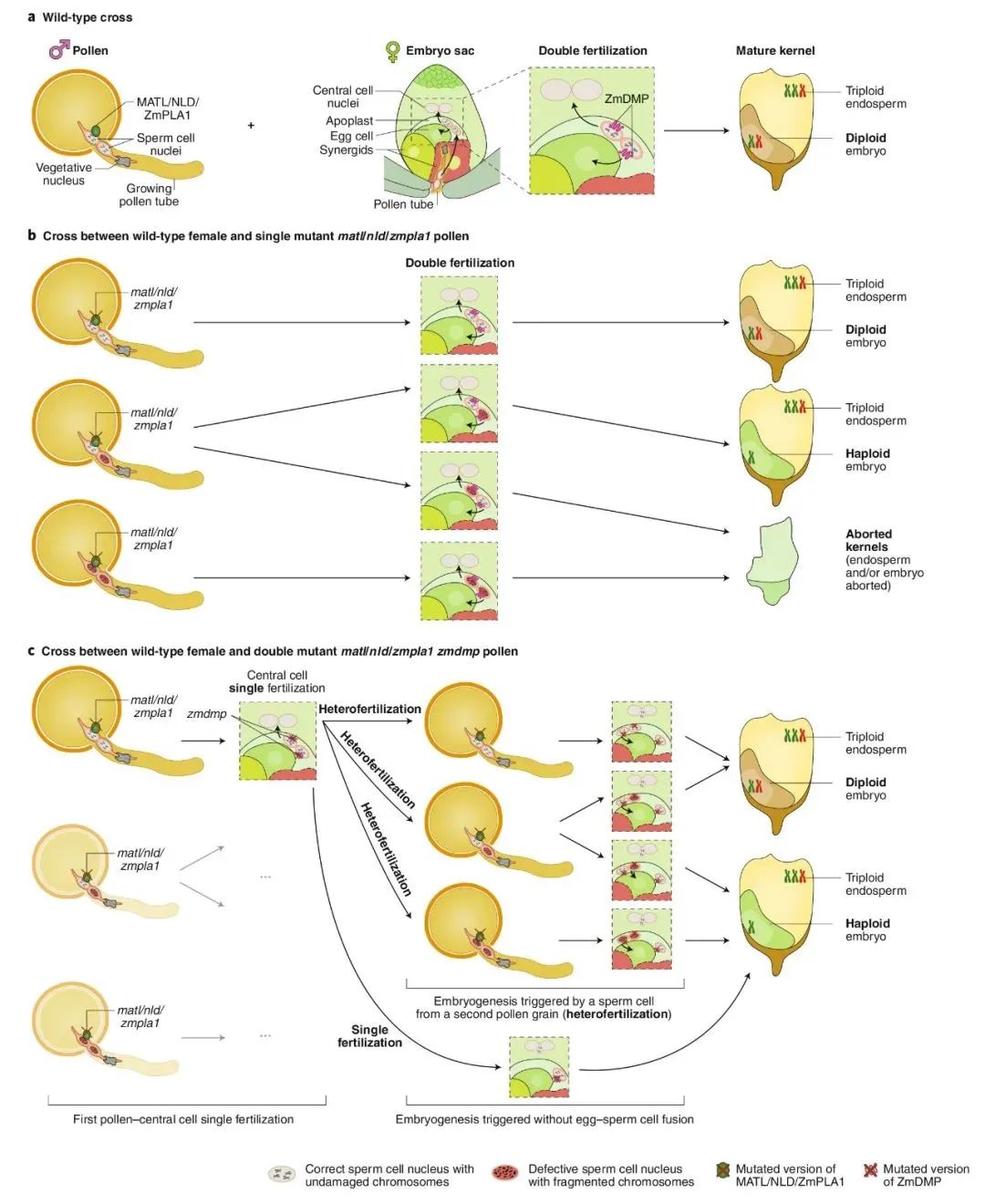
图2 玉米单倍体诱导机制模型(Jacquier et al., 2020)。(a)玉米种子发育是由双受精触发的,在双受精过程中,单倍体卵细胞和二倍体中央细胞分别与精子细胞融合,产生二倍体胚胎和三倍体胚乳;(b)matl/nld/zmpla1突变体与野生型杂交时,有三种类型的花粉粒,分别含有零个、一个或两个有缺陷的精细胞。因此,双受精后生成正常的籽粒、具有单倍体胚的籽粒或败育的籽粒;(c)与matl/nld/zmpla1 zmdmp双突变体花粉杂交的筛选结果表明zmdmp突变对单倍体诱导的促进作用。
着丝粒组蛋白3基因(CENH3)是近几年报道的创制单倍体诱导系的关键基因。CENH3仅存在于真核生物的功能着丝粒中,其主要参与着丝粒复合蛋白的募集和稳定,是细胞分裂时染色体分离所必需的,对着丝粒在染色体上的定位起重要作用(Sullivan et al., 2001)。类似于其他组蛋白,CENH3由两个结构域组成,一个N端尾巴(从核小体伸出,是翻译后表观修饰的靶点)和一个C端组蛋白折叠结构域(c-terminal histone fold domain, HFD)(Britt and Kuppu, 2016)。CENH3在植物中广泛存在,并且其功能在各个物种中保守。以CENH3表达发生改变的基因型植株为母本与野生型杂交,可使其基因组在合子中消失,而只保留野生型基因组的单倍体植株(图3)。CENH3介导的单倍体诱导方法目前已经在拟南芥、玉米、小麦上取得了重要的进展,未来还将扩展到更多的植物中。有关这一部分的内容,小远在先前的推文中有详细的讲解,大家感兴趣的话可以查阅:纯系养成——单倍体诱导技术(二)。
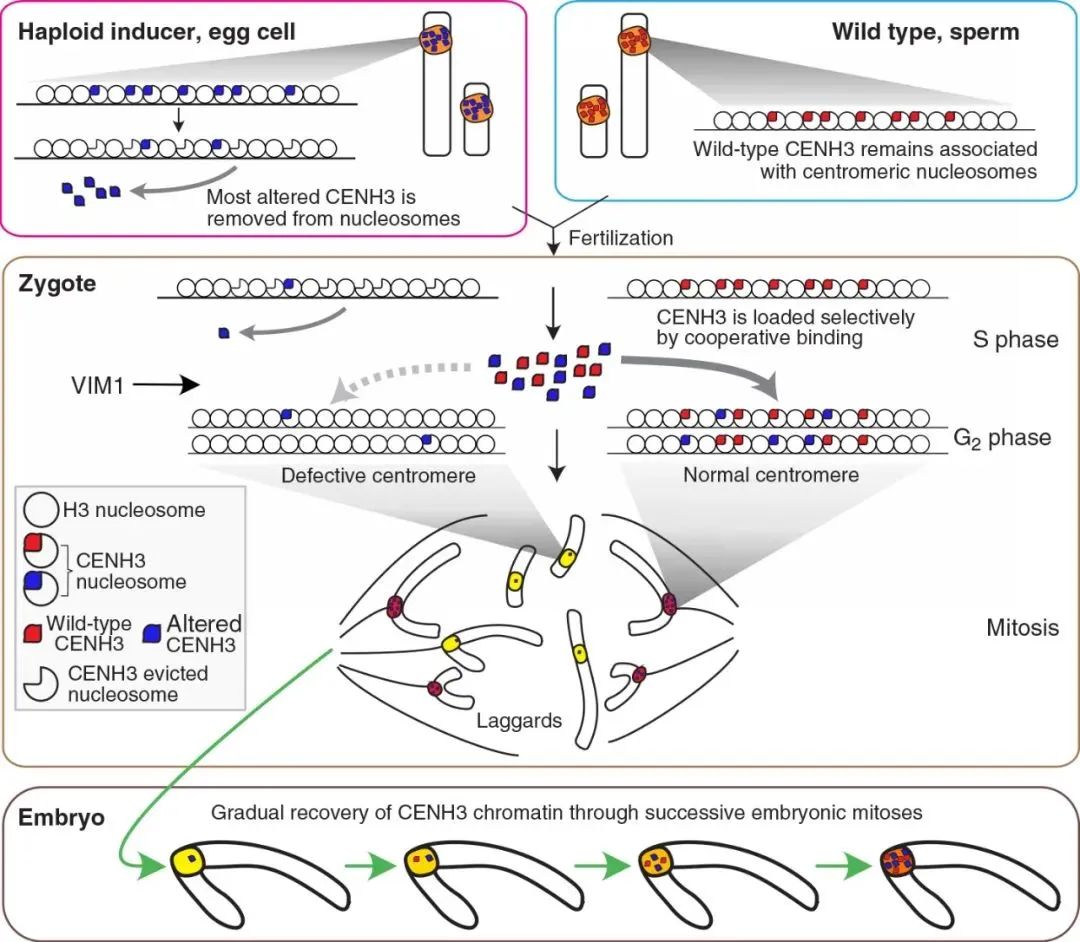
图3 拟南芥中CENH3介导的单倍体诱导模型(Marimuthu et al., 2021)。在成熟的母细胞中,CENH3变体会被选择性地从染色质中去除,但野生型中的CENH3不受影响;杂交后,CENH3弱化的母本着丝粒会与父本野生型着丝粒竞争加载CENH3;由于协同结合效应,CENH3会优先加载在野生型亲本的着丝粒上;在随后的有丝分裂过程中,单倍体诱导系的染色体由于其弱着丝粒而不能正常分离;其中,VIM1介导的泛素化或甲基化可以影响CENH3的稳定性,并且VIM1有利于CENH3的加载。
单倍体诱导之后,从众多杂交种中简便快速的筛选出单倍体籽粒是单倍体育种工程化的重要环节之一。目前单倍体鉴别的方法有很多,常用的方法包括:植株形态、细胞遗传学、遗传标记鉴别等方法。
2.1 利用植株形态鉴别单倍体
鉴别单倍体最基础的方法就是形态鉴别法。由于单倍体仅含有一组染色体,表现为细胞体积较小、植株的叶片面积、花茎以及种子等营养器官相较于二倍体小,植株也较矮(图4、图5、图6)。因此,通过观察植株的表型可以很直观的分辨出单倍体。但是该方法鉴定单倍体所需的时间较长,植物的生长也容易受到环境的影响造成鉴别失误,并且在农作物单倍体鉴定过程中需要在田间种植,因此容易造成耕地的浪费。

图4 将单倍体(Haploid)与二倍体(Diploid)玉米进行表型比较(Wang et al., 2021)。(A)单倍体植物株型矮小;(B)单倍体植株不育,没有花药。

图5 将单倍体(H)与六倍体(HE)小麦进行表型比较,包括株型、叶、穗和花药(Liu et al., 2020)。

图6 烟草单倍体(Haploid)和二倍体(Diploid)植株的表型比较(zhang et al., 2022)。(a)植物的整株表型;(b)烟草绽放的花;(c)未开放的花;(d)雌蕊;(e)雄蕊;(f)花药;(g)果荚。
2.2 结合细胞遗传学鉴别单倍体
通过染色体压片的方法观察植物的根尖细胞或减数分裂细胞是鉴别单倍体的最直观方法,通过观察染色体数是否等于配子数即可判断植株是否为单倍体。但该方法所涉及的实验步骤繁琐,且效率很低,不适用于工程化的筛选鉴定。流式细胞仪(Flowcytometry,简称FCM)是现代分子生物实验室不可或缺的高科技细胞分析仪器。它在检测细胞的大小,内部结构、DNA含量、特异性蛋白含量等方面具有显著优势,因此也被广泛应用于植物遗传学、植物生理学等多个领域(姚丽萍等, 2009)。利用流式细胞仪对大量处于分裂期的植物细胞DNA含量进行检测,经过计算机软件系统自动统计分析后,就能形成非常直观的DNA含量的分布曲线图来确定样本的倍性(图7)。使用流式细胞仪来检测单倍体的方法快速、稳定、直观、可靠,但是该仪器非常昂贵,并且需要专业的仪器维护人员和专业熟练的操作才能得到真实可靠的结果,这制约了该项技术的应用范围。
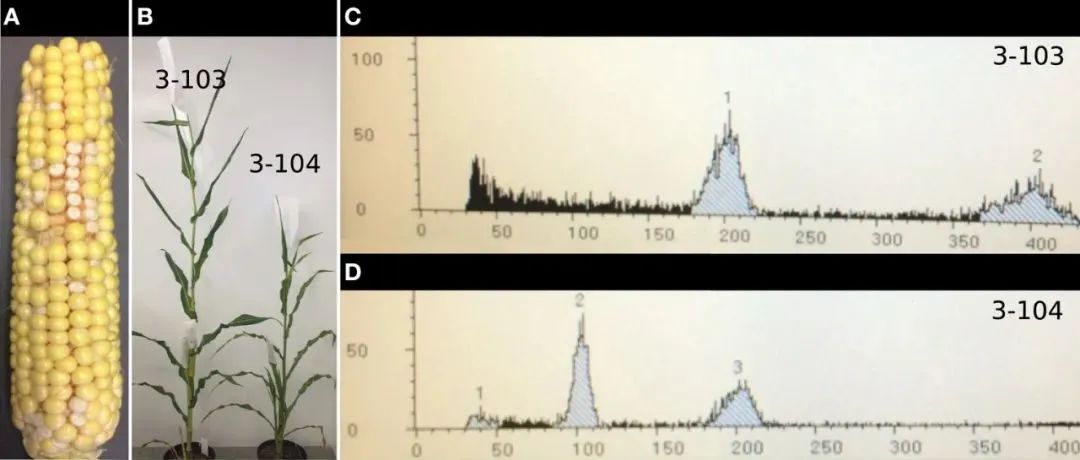
图7 用流式细胞仪检测玉米单倍体(Kelliher et al., 2016)。
2.3 利用遗传标记鉴别单倍体
(1)R1-nj 标记
目前各大育种公司和科研院所使用的单倍体遗传标记大多是基于R1-nj基因的遗传标记法。该基因可以在玉米胚乳的糊粉层和胚的盾片处合成花青素,通过肉眼就可以观察到明显的紫色。使用含有该遗传标记的单倍体诱导系,获得的单倍体籽粒表现为胚部不着色,胚乳部分着色,因此很容易从玉米籽粒中筛选出来。但是,R1-nj的效率受遗传背景和环境因素的影响较大,识别准确率在60%到90%之间(图8)。因此,这种方法在大规模筛选单倍体时存在一定的劣势。
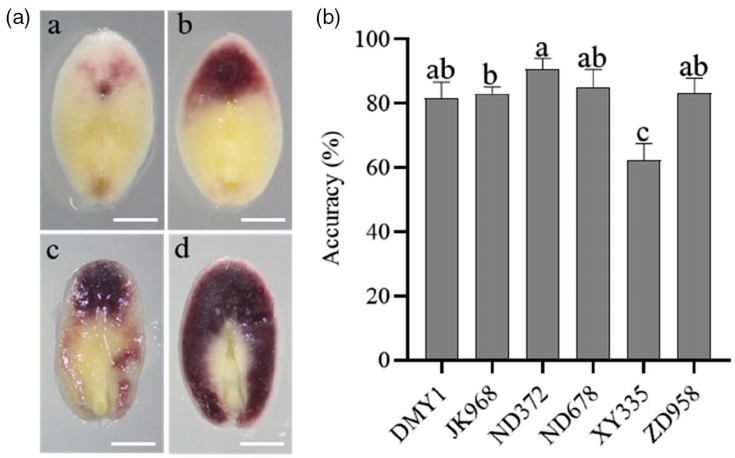
图8 不同杂交种中R1-nj色素沉着情况及单倍体识别的准确性(Chen et al., 2022)。
(2)MAGIC系统
MAGIC系统是近段时间陈绍江团队和陈茹梅团队共同利用优化的花青素标记物开发出的新型单倍体识别系统。与R1-nj标记相比,MAGIC1在鉴别单倍体方面的准确率达到了99.1%,并且在授粉后12天就可以鉴别单倍体,因此更适合大规模的单倍体筛选。
该团队在开发MAGIC系统之初,发现转录因子ZmC1和ZmR2的共表达足以使玉米产生紫色愈伤。通过瞬时转化分析发现,ZmC1和ZmR2共表达后在玉米胚和籽粒糊粉层中均有花青素产生。将含有ZmC1和ZmR2的稳定转基因株系与自交系杂交后,其后代也全部积累了花青素。这些结果均表明ZmC1和ZmR2可以作为稳定的遗传标记物。因此,他们将CAU6作为亲本与上述稳定转基因株系进行回交后,再进行四代自花授粉,最终筛选到了含有稳定遗传标记的单倍体诱导系MAGIC1。随后,该团队将含有ZmC1和ZmR2的表达载体中的启动子优化成胚胎特异性和种子特异性双向启动子,获得了单倍体鉴别率以及种子发芽率更高的升级版MAGIC2(图9)。
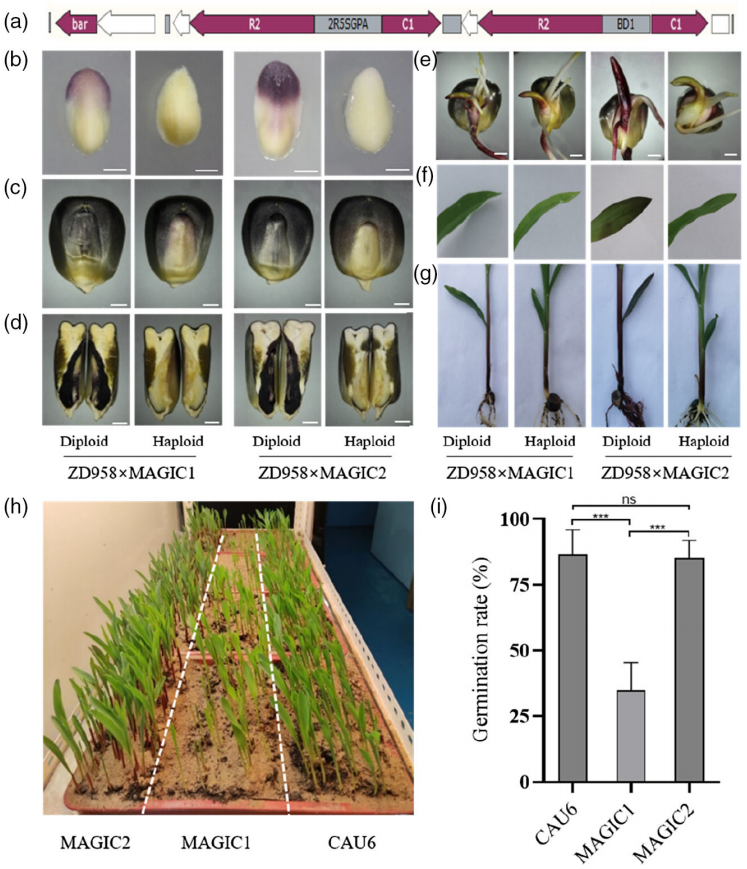
图9 MAGIC1和MAGIC2花青素积累和发芽率差异(Chen et al., 2022)。(a)p2Seed-C1R2载体的示意图,2R5SGPA:种子特异性双向启动子;(b)15DAP的未成熟胚胎;(c)籽粒;(d)籽粒的垂直切面;(e)胚芽鞘;(f)叶子;(g)鞘和根;(h)三种单倍体诱导系的发芽率表现;(i)三种单倍体诱导系的发芽率柱状图。
(3)高油性状标记
利用高油型诱导系与普通玉米杂交获得的杂合二倍体和单倍体子粒油分差异大。因此,采用核磁共振仪器或近红外仪器对单倍体籽粒进行检测,相较于人工鉴别会大大缩短检测时间(Li et al., 2009)。而进一步提高油型诱导系的诱导率和油分含量,不仅能够提高单倍体诱导效率,还能提高鉴别的效率和准确率,从而实现工程化的单倍体育种。陈绍江团队利用单倍体诱导系CAU2和北农大高油群体(BHO)作为供体亲本,培育出了新型高油高效玉米单倍体诱导系CHOI4,其平均单倍体诱导率为15.78%,籽粒平均油分含量达11%,这一成果大幅提高了单倍体的鉴别效率(图10)。
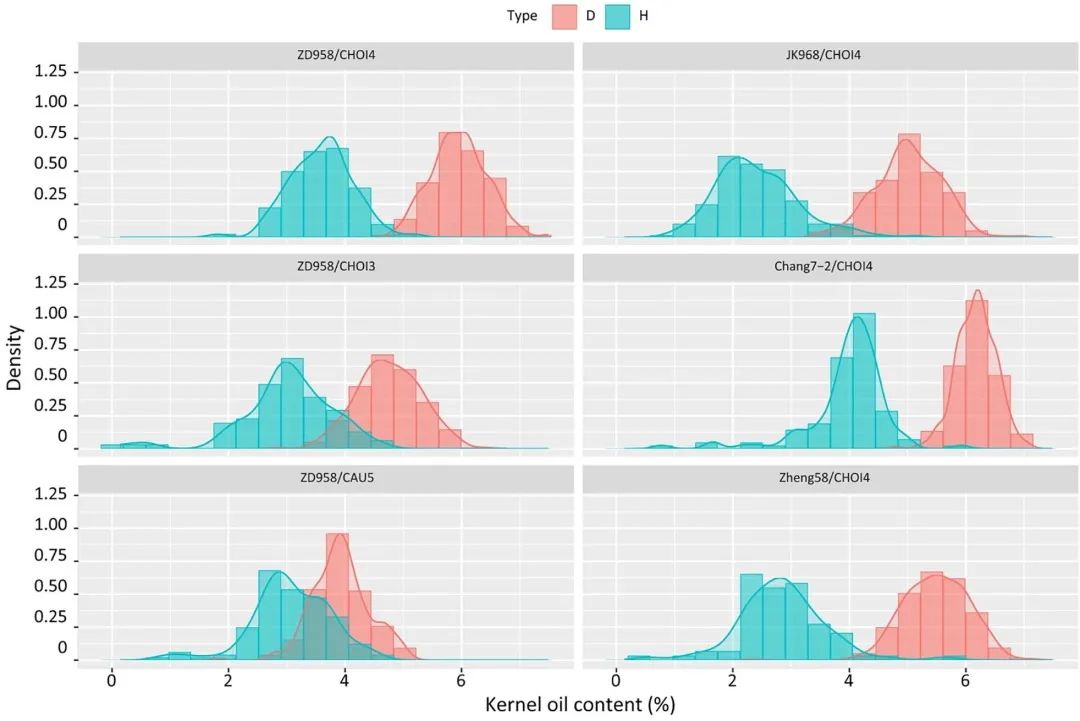
图10 来自不同杂交组合的单倍体(H)和二倍体(D)的含油量(KOC)比较(Liu et al., 2021)。左侧:来自ZD958×CHOI4(上)、ZD958×CHOI3(中)和ZD958×CAU5(下)的二倍体(粉红色)和单倍体(浅蓝色)的含油量。右侧:分别来自JK968×CHOI4、Zheng58×CHOI4和Chang7-2×CHOI4杂交的单倍体和二倍体种子的含油量。
(4)荧光蛋白标记
将含有荧光标记(YFP、GFP等)的转基因植株与创制的单倍体诱导系进行杂交,得到含有稳定荧光标记的单倍体诱导系后。这种诱导系获得的杂交种很容易借助荧光体式显微镜、激光共聚焦显微镜等荧光观察工具,来对其中的单倍体籽粒进行鉴别筛选。例如,谢传晓等人利用胚特异型启动子ZmESP和胚乳特异性启动子HvASP,构建了含有胚特异表达绿色荧光蛋白和胚乳特异表达红色荧光蛋白的双荧光标记单倍体诱导系,将其作为父本与三个受体品种ZD99、Zeyu7、Feng8进行杂交。通过相应的滤光片在不同波长的激发光下,可以观察到在二倍体和单倍体籽粒中的红色胚乳,而仅在二倍体籽粒中观察到绿色胚(图11F)。陈绍江团队为解决番茄单倍体的筛选难题,也建立起了基于FAST-Red的荧光快速鉴别方法,通过浸湿种子和刚萌发根尖荧光的两步观察,可实现番茄单倍体的快速精准鉴别(图12)。在花青素标记或油份含量筛选标记因遗传背景受到限制时,荧光标记法仍能实现快速、高效的单倍体籽粒鉴别。
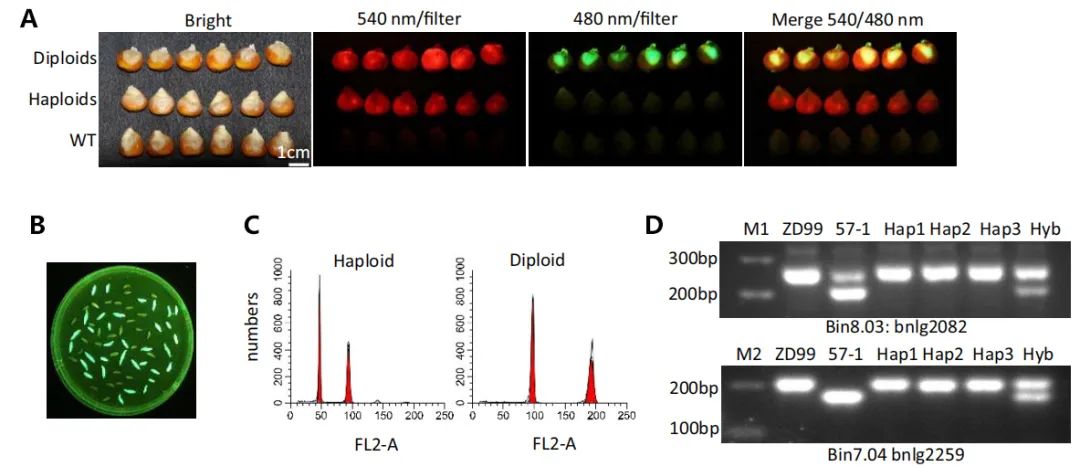
图11 玉米中基于荧光蛋白标记的高效单倍体鉴别系统(Dong et al., 2018)。(A)不同激发光条件下二倍体和单倍体籽粒的差别。(B)480nm滤光片下未成熟的二倍体胚(较大的带有eGFP荧光)和单倍体胚。(C)使用流式细胞仪对单倍体(左)与二倍体(右)的DNA含量进行验证。(D)使用SSR标记检测的电泳结果图。
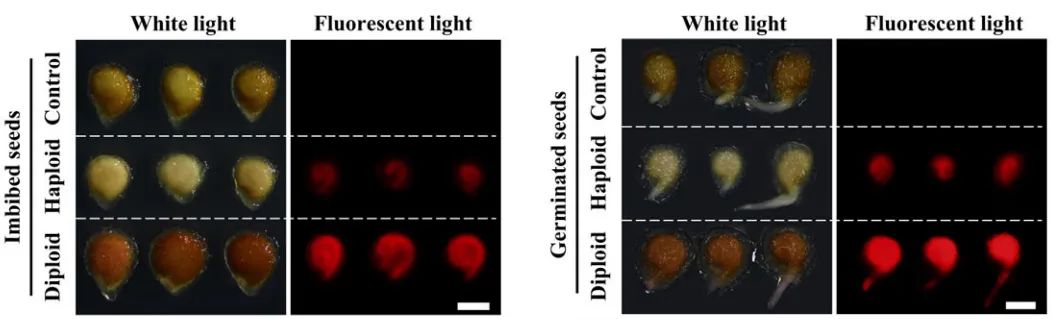
图12 基于FAST-Red的单倍体种子鉴定(Zhong et al., 2021)。胚乳中RFP表达较弱且胚中没有RFP表达的白色种子被认定为假定的母体单倍体。
2.4 单倍体加倍
经过单倍体鉴别后,就可以进入单倍体育种的关键环节—单倍体加倍。虽然在没有人为干预的条件下,单倍体就可以完成自然加倍,但是因为加倍效率较低,无法满足种业公司的育种要求。因此,目前常用的玉米单倍体加倍方法是化学加倍法,常以秋水仙素作为加倍剂来对单倍体进行加倍(葛百一等,2018)。利用秋水仙素来对单倍体进行加倍的方法主要包括:浸种法、浸芽法、浸根法、滴心法、注射法和组织培养法。
(1)浸种法
浸种法操作简单,是诱导后比较常用的加倍方法。先用清水浸泡种子(24h),再用不同浓度梯度秋水仙素溶液进行处理,最后经流水冲洗一段时间,确保残留的秋水仙素洗净后进行播种即可。其加倍效率易受到秋水仙素浓度、浸泡时间的影响。
(2)浸根法
浸根法是指用秋水仙素浸泡单倍体植株根部的方法。将幼苗期的植株从土壤中拔出并洗净根部的泥土后,将其浸泡在秋水仙素溶液中一段时间,然后用流水洗净根部残留的秋水仙素,再将其种植于大田中。该方法耗费的秋水仙素剂量大,成本较高,同时涉及到育苗、处理、移栽等多个环节,幼苗会受到毒性和移栽损伤的双重影响,因此其成活率较低。
(3)浸芽法
浸芽法是指用秋水仙素浸泡幼芽的方法。当单倍体种子幼芽生长至2cm左右时,切掉部分顶端,将幼芽浸泡于秋水仙素溶液中一段时间,再移栽到育苗盘中,当幼苗长出4-5片叶子时可移栽到大田。由于该方法将整个幼芽浸泡在秋水仙素溶液中,幼芽吸收了大量的秋水仙素溶液,缓苗所需的时间较长,并且容易造成大量的幼苗死亡。
(4)滴心法
滴注心叶法是将单倍体幼苗移栽于营养皿后,将配比好的秋水仙素溶液滴注于三叶期或者五叶玉米单倍体幼苗的心叶内,经过缓苗处理后可移栽大田进行精细管理。
(5)注射法
注射法采用微量注射器,将一定浓度的秋水仙素溶液注入植株顶芽或侧芽中,来完成单倍体的加倍。注射法不需要育苗、移栽,操作简便且用量少,但需要对田间每一单倍体植株进行注射,劳动强度较大,由于田间植株长势不一致,对注射的技术要求很高。
(6)组培法
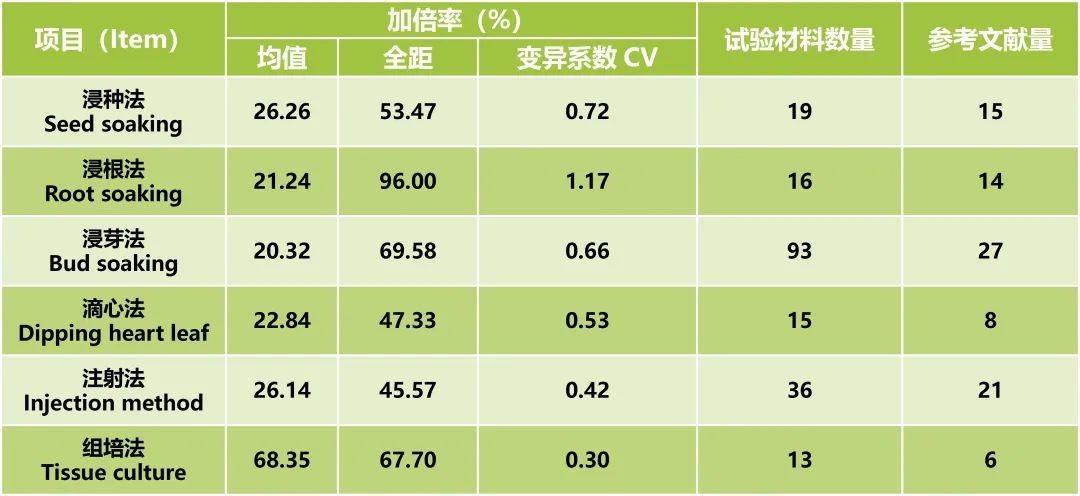
组织培养加倍是通过培养基渗入的途径,对单倍体幼胚等愈伤组织进行加倍处理。由于实验条件控制精确,使得实验结果重复性强。对国内外研究中利用秋水仙素对玉米加倍的不同方法进行统计发现,组培法的加倍率最高(表1)。但由于实验环境要求较高,操作程序复杂,使得组培加倍法在单倍体育种中的应用受到一定的限制。
表1 秋水仙素不同加倍方法加倍率统计(郭书磊等, 2020)。
单倍体育种因具备周期短、纯度高等优点,获得了国内外各大农业公司及育种单位的密切关注。对于种业公司来说,早日推出优异新品种就可以早日获取效益,而对于育种家们来说,缩短育种周期、加快育种速度是他们毕生的奋斗目标。在过去的几十年里,单倍体育种技术已经逐步成为玉米育种上能够规模化应用的关键核心技术。近几年,国内外玉米育种企业和农业科研院所使用单倍体育种技术创制的纯系所组配的杂交种数量迅速提升(图13)。2020年,国内36家科研单位和育种企业通过审定的单双倍体杂交种数量已累计达到165个,并且,随着单倍体育种技术应用规模的不断扩大,越来越多的玉米种子企业实现了工厂化的大规模应用,此外专业进行玉米单倍体加倍的公司也陆续出现,年生产的加倍单倍体苗达到上千万株。其它农作物或经济作物的单倍体育种虽然没有玉米单倍体育种的应用程度高,但是随着技术的不断突破,工厂化应用将很快到来。
图13 单倍体育种技术的应用现状(图片参考刘晨旭博士PPT)。
文章至此就告一段落了,随着单倍体诱导关键调控基因的进一步挖掘和基因编辑技术的联合使用,单倍体育种技术已经不仅仅局限在玉米纯系的创制上,其应用也由玉米拓展到单子叶作物水稻、小麦、谷子,以及双子叶拟南芥、蒺藜苜蓿、番茄、烟草等多种植物上,未来单倍体育种技术在作物育种和改良上将发挥更大的作用,粮食作物以及蔬菜经济作物实行工厂化育种指日可待!最后,为大家简单总结一下基于种内杂交的单倍体育种思路,一起来看看吧~
高效单倍体育种思路