
图 1. APOE4 基因是 AD 的主要遗传风险因素[3]
a:三种 APOE 蛋白突变体的结构;b:APOE4 基因剂量依赖性增加 AD 风险并降低发病年龄
■ APOE4,阿尔兹海默症 “预判者”?
作为阿尔茨海默病的主要易感基因,APOE4 如何做到“预判你的预判”?血脑屏障 (BBB) 的破坏是人类认知功能障碍的早期生物标记物。2020 年 4 月,Montagne 等人发表在 Nature 的研究结果证明:APOE4 相关的认知衰退与血脑屏障 (BBB) 损伤有关,并且这种损伤早在 AD 发生之前就可以发生。
作者团队发现认知正常者的 APOE4 (ε3/ε4 和 ε4/ε4) 携带者的海马 (HC) 和内侧颞叶中 (PHG) 中的血脑屏障被破坏,但临床痴呆评分 (CDR) 评分均为0。当 CDR=0.5 时,APOE4 携带者 HC 和 PHG 的血脑屏障破坏会进一步增加,且这种增加与脑脊液 (CSF) Aβ 和 tau 蛋白变化无关。

图 2. APOE4 携带者海马和海马旁回血脑屏障的破坏[5]
a.HG 和 PHG 的示意图 b. 动态对比增强磁共振成像技术在 APOE3 纯合子 和 APOE4 携带者的海马区 (HC) 生成血脑屏障 (BBB) 通透性 Ktrans 图。c. CDR 0 或 0.5 的情况下, APOE4 携带者 HC 和 PHG 中的 Ktrans 值。
此外,Montagne 等人还研究发现与 APOE3 携带者相比,APOE4 携带者的脑脊液中周皮细胞损伤的生物标志物: 一种可溶性的蛋白质称为血小板源生长因子受体-β (sPDGFRβ) 也会升高。
AD 进程中,APOE 如何发力?
■ APOE 同种型的不同效力
ApoE2, ApoE3, and ApoE4 Differentially Stimulate APP Transcription and Aβ Secretion 一文表明人类神经胶质细胞 ApoE 的三种遗传同种型 (ApoE2、ApoE3 和 ApoE4) 激活 APP 基因转录和 Aβ 合成方面的表现出不同的效力。
ApoE 以 ApoE4 > ApoE3 > ApoE2 效力等级顺序激活非经典 MAP 激酶信号通路: DLK → MKK7 → ERK1/2 MAP 激酶途径,并且增强了 APP 转录和淀粉样蛋白-β 合成。

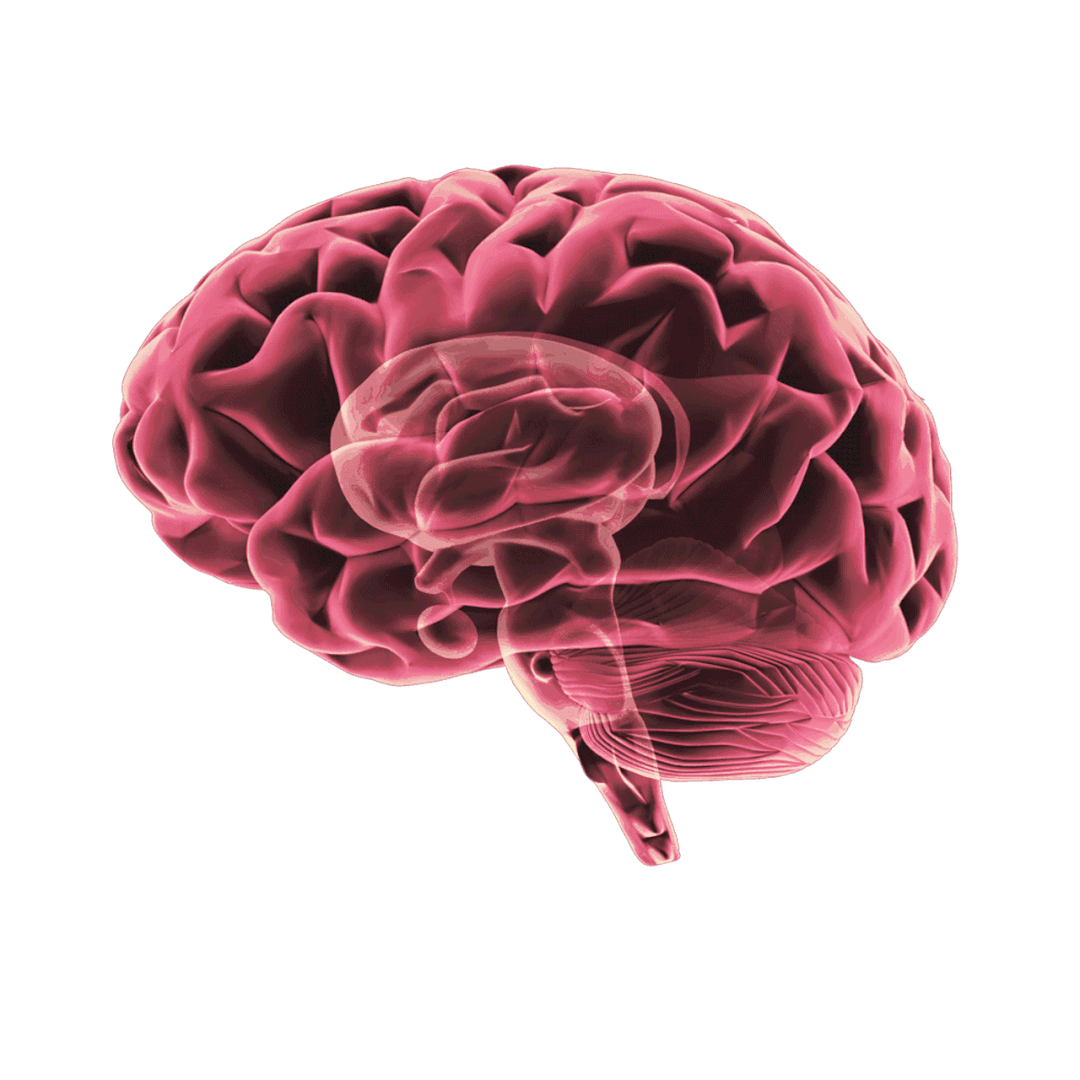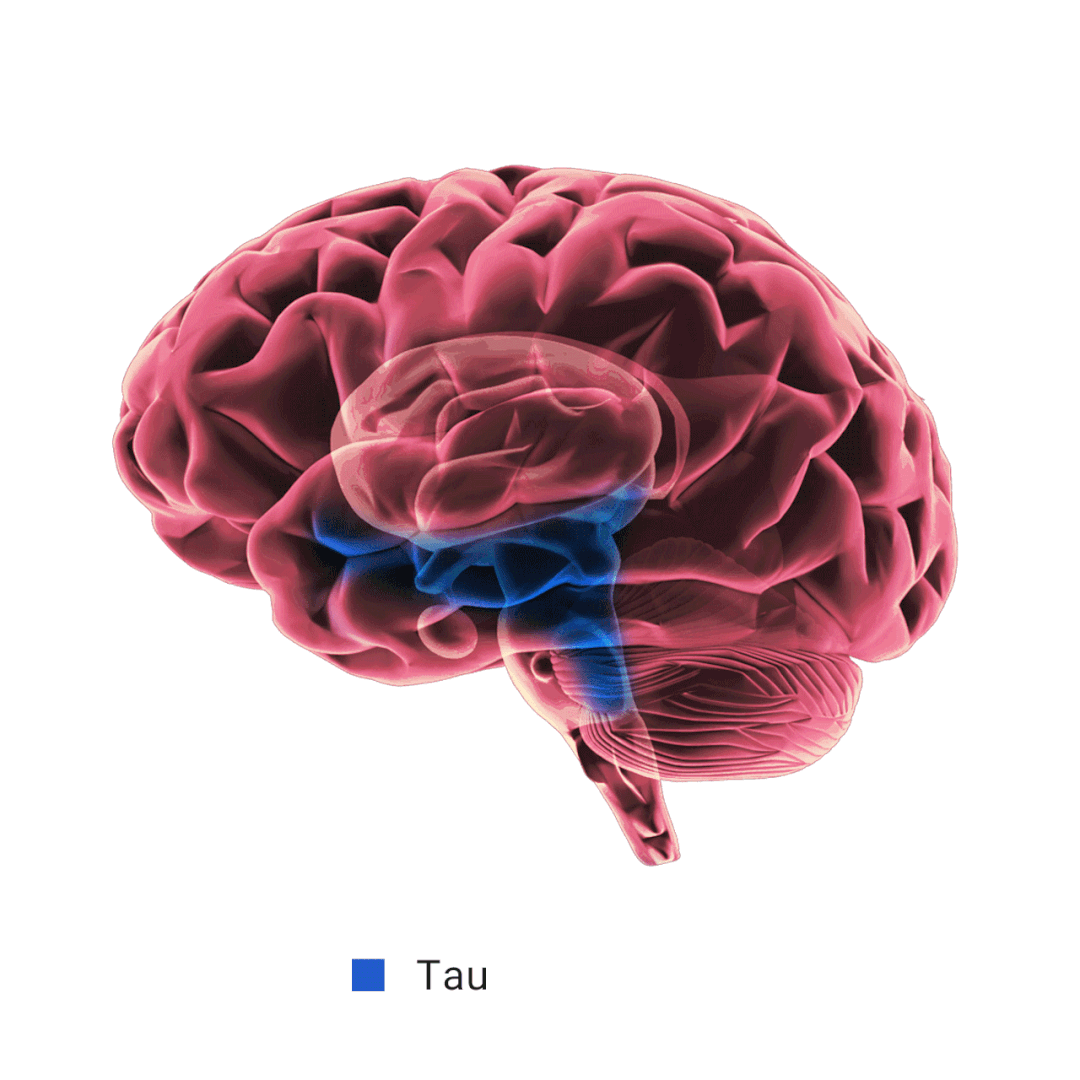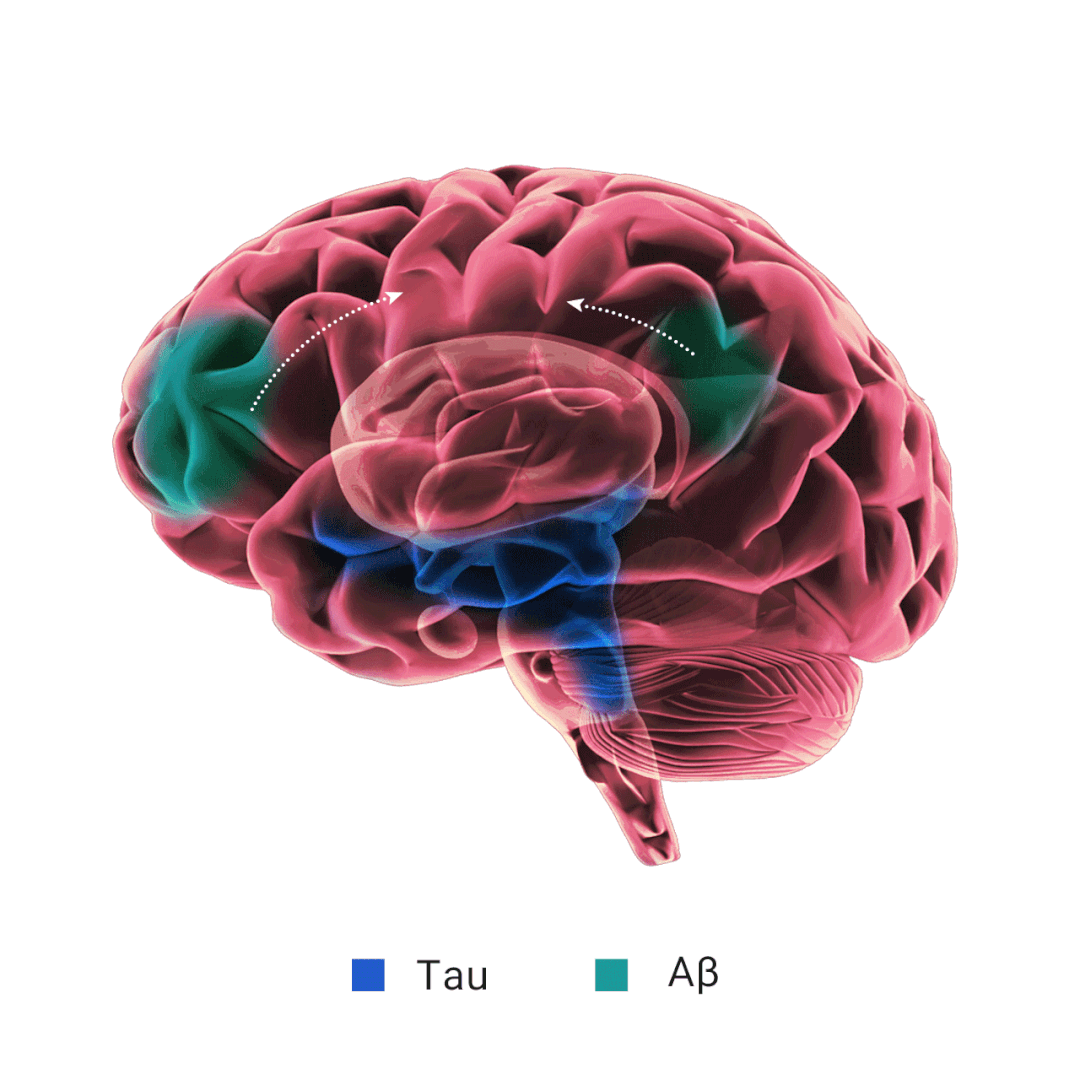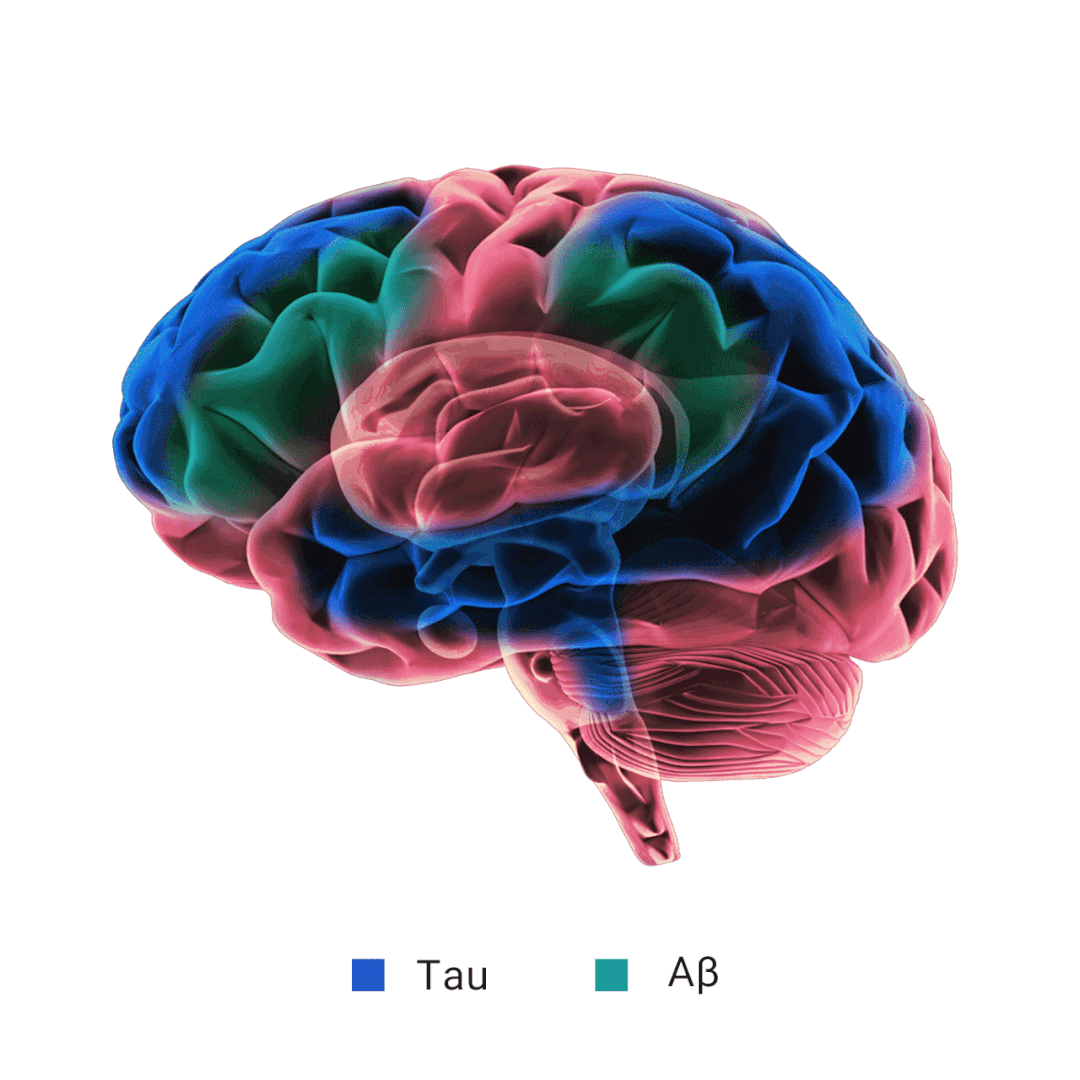
当 Aβ 产生过多和/或清除效率不高时,就会发生 Aβ 聚集。淀粉样斑块的主要成分 Aβ 纤维的形成经历三个动力学阶段: 滞后期、生长期和平台期。APOE 对 Aβ 纤维形成的影响不一致,研究表明,APOE 在 Aβ 沉积的初始播种阶段起着至关重要的作用,在这个时期星形胶质细胞 APOE4 过表达会加剧淀粉样病变。

图 4. APOE 的异构体和 Aβ 的聚集和清除[9]
a. Aβ 从大脑中被移除的主要途径包括血脑屏障的清除、细胞摄取和随后的降解、酶降解、ISF体流的清除以及脑脊液吸收进入循环和淋巴系统。b. 大脑 Aβ 清除不足导致Aβ 积累。
■ APOE4 如何损伤大脑?
除了上述与“主流”观点相关的 APOE4 影响 AD 进程论。Thomas E. Willnow 等人在 Alzheimers Dement 杂志上发表了一项新研究,探索了AD 进程中,ApoE4 基因损伤大脑的新机制。
APOE 作为一种脂质转运蛋白,可以为神经元提供重要营养物质,包括不饱和脂肪酸 (神经元周围细胞膜的组成部分,PUFA)。APOE 的结合受体 sortilin,该受体介导将结合 apoE 的货物 (apoE-bound cargo;也就是营养成分) 摄取到神经元中。
作者团队将 Sort1 defect (KO) 引入到带有编码人类 APOE3 或 APOE4 基因座,靶向替换小鼠 Apoe 位点 (E3; KO 或者 E4; KO)。与野生型鼠 (WT) 相比,sortilin 的丢失会导致 KO 小鼠的皮质和海马中人 APOE3 和 4 (图 5a)。在 sortilin 缺陷小鼠中, Aβ40 水平增加了两倍 (图 5b)。

APOE4 破坏了 Sortilin 的营养物质摄取作用[12]
a. APOE3/4 的脑积聚。b. 可溶性 Aβ40 沉积。c.多不饱和脂肪酸 (PUFA) 的水平变化。d-e, APOE4 破坏了 sortilin 的细胞表面再循环
作者团队还使用基于 LC-MS 的脂质组学来确定 (E3;KO) 和 (E4;KO) 小鼠脑皮质中各种不饱和脂肪酸 (PUFA) 的水平。APOE3 小鼠中,sortilin 的缺失会损害神经保护性 PUFA 摄取途径,增加淀粉样蛋白形成。APOE4 的野生型小鼠中有类似的现象。APOE4 会损害 sortilin 介导的神经元对脂质的摄取途径(图 5d-e)。这种损害使得神经细胞吸收的脂肪酸越来越少,神经细胞无法保护自己,导致发炎。
靶向 APOE 的治疗
在 AD 领域,靶向 APOE 的治疗主要集中在以下几个方面。
■ 增加 APOE 水平及其脂质化
大脑中的 APOE4 比 APOE2 和 APOE3 的脂化水平低且稳定,因此,增加大脑中的 APOE4 与脂质的结合,可能加速 Aβ 的清除。Bexarotene 通过诱导 ABCA1 和 ABCG1 基因表达,增强 APOE 的脂质化,并增加 APOE 的表达水平,可快速降低 AD 小鼠脑内的 Aβ 斑块负荷,改善认知能力。目前 Bexarotene 正在进行针对于 AD 治疗的临床试验 (NCT01782742)。
■ 阻断 APOE 与 Aβ 的相互作用
抗 APOE 单克隆抗体 (HJ6.3) 可显著降低 APP/PS1 转基因记忆缺陷小鼠脑内不溶性 Aβ 水平、Aβ 斑块负荷和 APOE 水平。利用一种与 APOE 结合的合成多肽 Aβ12-28P 干扰 APOE 和 Aβ 的结合,可降低记忆缺陷小鼠的可溶性和不溶性 Aβ 水平和 Aβ 斑块负荷,并减少了 APOE 在 Aβ 斑块中的沉积。
■ APOE 类似物
APOE 类似物 (APOE 的 N 端片段,包括其受体结合基序) 已被证明可改善各种阿尔茨海默病小鼠模型中的 Aβ 水平和 Aβ 斑块负荷、tau 过度磷酸化和神经炎症,但尚未进入人类临床试验。
■ 基因治疗
CRISPR-Cas9 编辑技术用于编辑转换 APOE 等位基因的应用已在细胞培养层面获得成功,但仍有待在小鼠敲除实验中验证。然而,应用基因治疗来表达 APOE ε2 并提高 APOE ε4 携带者 (甚至是 APOE ε3 纯合子) 中的 APOE ε2 表达水平已成为可能,该方法正在进行临床试验 (NCT0363407)。
不难看出,科学家们对于 APOE 在 AD 方面的研究目前主要还停留在动物甚至是细胞或分子层面,真正进入临床试验的凤毛麟角,但是该靶点仍然被寄予厚望,希望在不久的将来能看到新的突破。

MCE 的所有产品仅用作科学研究或药证申报,我们不为任何个人用途提供产品和服务。
参考文献
1. Strittmatter WJ, Roses AD. Apolipoprotein E and Alzheimer's disease. Annu Rev Neurosci. 1996;19:53-77.
2. Genome-wide association study identifies variants at CLU and PICALM associated with Alzheimer's disease. Nat Genet. 2009 Oct;41(10):1088-93..
3. Liu CC, Liu CC, Kanekiyo T, Xu H, Bu G. Apolipoprotein E and Alzheimer disease;risk,mechanisms and therapy. Nat Rev Neurol.2013 Feb;9(2):106-18.
4. Mahley RW. Apolipoprotein E: cholesterol transport protein with expanding role in cell biology. Science. 1988 Apr 29;240(4852):622-30.
5. Axel Montagne, Daniel A Nation, et al. APOE4 leads to blood-brain barrier dysfunction predicting cognitive decline. Nature. 2020 May;581(7806):71-76.
6. Wahrle SE, Jiang H, Parsadanian M, Legleiter J, Han X, Fryer JD, Kowalewski T, Holtzman DM. ABCA1 is required for normal central nervous system ApoE levels and for lipidation of astrocyte-secreted apoE. J Biol Chem.2004 Sep 24;279(39):40987-93.
7. Karten B, Campenot RB, Vance DE, Vance JE. Expression of ABCG1, but not ABCA1, correlates with cholesterol release by cerebellar astroglia. J Biol Chem. 2006 Feb 17;281(7):4049-57.
8. Yu-Wen Alvin Huang, Bo Zhou, Marius Wernig, Thomas C Südhof, et al. ApoE2, ApoE3, and ApoE4 Differentially Stimulate APP Transcription and Aβ Secretion. Cell. 2017 Jan 26;168(3):427-441.e21.
9. Rebeck GW, Reiter JS, Strickland DK, Hyman BT. Apolipoprotein E in sporadic Alzheimer's disease: allelic variation and receptor interactions. Neuron. 1993 Oct;11(4):575-80.
10. Huang YA, Zhou B, Wernig M, Südhof TC. ApoE2, ApoE3, and ApoE4 Differentially Stimulate APP Transcription and Aβ Secretion. Cell. 2017 Jan 26;168(3):427-441.e21.
11. Deane R, Sagare A, Hamm K, Parisi M, Lane S, Finn MB, Holtzman DM, Zlokovic BV. apoE isoform-specific disruption of amyloid beta peptide clearance from mouse brain. J Clin Invest. 2008 Dec;118(12):4002-13.
12. Antonino Asaro, Anne-Sophie Carlo-Spiewok, Anna R Malik, et al. Apolipoprotein E4 disrupts the neuroprotective action of sortilin in neuronal lipid metabolism and endocannabinoid signaling. Alzheimers Dement. 2020 Sep;16(9):1248-1258.
13. Ma Q, Zhao Z, Sagare AP, Wu Y, Wang M, Owens NC, Verghese PB, Herz J, Holtzman DM, Zlokovic BV. Blood-brain barrier-associated pericytes internalize and clear aggregated amyloid-β42 by LRP1-dependent apolipoprotein E isoform-specific mechanism. Mol Neurodegener. 2018 Oct 19;13(1):57.
14. Hawkes CA, Sullivan PM, Hands S, Weller RO, Nicoll JA, Carare RO. Disruption of arterial perivascular drainage of amyloid-β from the brains of mice expressing the human APOE ε4 allele. PLoS One. 2012;7(7):e41636.
15. Shi Y, Yamada K, et al. ApoE4 markedly exacerbates tau-mediated neurodegeneration in a mouse model of tauopathy. Nature. 2017 Sep 28;549(7673):523-527.
16. Serrano-Pozo A, Das S, Hyman BT. APOE and Alzheimer's disease: advances in genetics, pathophysiology, and therapeutic approaches. Lancet Neurol. 2021 Jan;20(1):68-80.Erratum in: Lancet Neurol. 2021 Feb;20(2):e2.

