

撰文︱王思珍
责编︱王思珍
AMPA全称α-氨基-3-羟基-5-甲基-4-异唑丙酸,NMDA全称N -甲基-D-天门冬氨酸。诸多研究表明,AMPA受体(AMPARs)和NMDA受体(NMDARs)在调节大脑兴奋性突触应答、控制突触特性、影响突触可塑性、以及在神经精神疾病发病中起到重要作用【1-3】。 然而,对于NMDAR和AMPAR调控突触组成的机制,知之甚少。
neurexins属于突触前细胞黏附蛋白家族,在突触连接神经元中起作用,与精神分裂症、自闭症和图雷特综合症有关【4-5】。(补充:图雷特综合症(Tourette’s syndrome)是一种频繁运动性抽动和声带抽动的遗传性疾病,症状只出现在白天,且持续1年以上)。 有报道称,在海马神经元突触中,neurexin-1的特异剪接变体(简称NRXN1SS4+)可以选择性地增强NMDAR介导兴奋性突触后电流,而相反地,neurexin-3的剪接变体(简称NRXN3SS4+)则抑制AMPAR介导兴奋性突触后电流【6-7】。然而,这两种剪接变体是如何差异调控AMPAR和NMDAR的,目前不清楚。
离子型谷氨酸Δ受体1和2(GluD1和GluD2)具有突触后离子型谷氨酸受体的分子结构,GluD1/2与分泌型小脑肽(CBLN1,CBLN2)结合而组装成跨突触黏附复合物,而小脑蛋白又与突触前neurexins相互作用【8-9】。然而,neurexins-小脑肽-GluD1/2复合体是否具有黏附性突触形成功能或介导跨突触信号?目前也不清楚。
近日,在以GluD1 is a signal transduction device disguised as an ionotropic receptor为题在线发表正在Nature上的最新论文中,斯坦福大学霍华德·休斯医学研究所的Thomas C. Südhof课题组(Jinye Dai(戴金叶)为文章第一作者和共同通讯作者)报道了NRXN1SS4+和NRXN3SS4+通过小脑肽依赖的GluDs激活来调节NMDARs和AMPARs, GluDs将不同的突触前轴突蛋白信号转化为不同的突触后受体AMPAR和NMDAR应答。

文章中,首先,体外体内实验发现,海马神经元中丰度较高的GluD1的缺失并不会损害突触形成,但会加强兴奋性突触后电流(简称EPSCs)的频率和振幅,也会提高AMPA/NMDA比率,且这种比率上升是由于AMPAR EPSCs振幅的增加(即分数的分子增大)和NMDAR-EPSCs振幅下降(即分数的分母减小),即突触后AMPAR和NMDAR的组成发生了改变(图1)。脑下脚(subiculum)小脑肽2(CBLN2)缺失也有类似现象(图2)。说明GluD1和CBLN2这两种调节机制作用于相同的突触,也暗示GluD1作为小脑肽-neurexins复合体的受体,从而发挥调节海马神经元NMDARs和AMPARs的功能,换言之,CBLN2作用于GluD1的上游。

图1 GluD1控制AMPARs和NMDARs而不改变突触形成
(图片引自:Dai, J. et al., Nature 2021; 595: 261–265)

图2 GluD1通过CBLN2依赖的机制控制突触后AMPAR EPSCs和NMDAR EPSCs
(图片引自:Dai, J. et al., Nature 2021; 595: 261–265)
紧接着的结果表明,GluD1介导了两种不同的轴突蛋白信号通路:NRXN1SS4+-CBLN2-GluD1-NMDAR和NRXN3βSS4+-CBLN2-GluD1-AMPAR。NRXN1βSS4+选择性地增强NMDAR,而非AMPAR,而NRXN3βSS4+则选择性地抑制AMPAR,但非NMDAR。GluD1缺失则会改信号通路,并产生相反的变化(图3)。而且,结果也表明,由于剪接位点(即SS4)所剪接的序列的特异性,导致了NRXN1SS4+和NRXN3SS4+调节突触后谷氨酸受体反应的特异性,提示neurexins通过与小脑肽结合从而控制AMPAR和NMDAR,表明小脑肽两种剪接体调控两种受体的信号特异性的起源为小脑肽的特异性剪接编码序列(图4a-k)。

图3 GluD1既能传导增强NMDAR EPSCs的NRXN1SS4+信号,又能传导抑制AMPAR EPSCs的NRXN3SS4+信号
(图片引自:Dai, J. et al., Nature 2021; 595: 261–265)
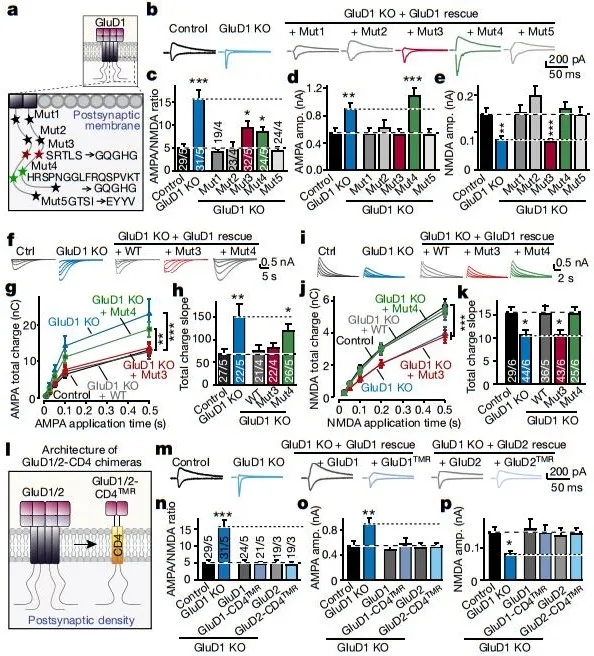
图4 不同的细胞质GluD1序列调节AMPARs和NMDARs,而不依赖于GluD1离子受体跨膜结构
(图片引自:Dai, J. et al., Nature 2021; 595: 261–265)
进一步地,作者发现,GluD1是通过不同的细胞质GluD1效应子序列(motif)差异性地将突触前NRXN1βSS4+和NRXN3βSS4+信号转导至突触后AMPAR和NMDAR应答。那么,GluD1是如何差异性地转导NRXN1SS4+和NRXN3SS4+信号的呢?即 GluD1信号转导机制是什么?作者推测,GluDs的离子受体结构通过构象变化从而介导信号传递,此外,GluDs可能直接将细胞外的neurexins-小脑肽信号转导到细胞内的特定响应以调节AMPARs和NMDARs。为此,作者对GluD1和GluD2不同结构域的进行了缺失和替换,发现,GluDs调节AMPARs和NMDARs的机制并不涉及其谷氨酸结合域或其离子受体跨膜结构(图4i-m,图5a-e)。

图5 短GluD1胞质序列基序将突触前neurexins信号转导为突触后AMPAR和NMDAR应答
(图片引自:Dai, J. et al., Nature 2021; 595: 261–265)
最后,作者探究了哪些GluD1最短(氨基酸序列)序列可能足以调节AMPARs和NMDAR,结果发现,含有细胞质序列基序3的最短序列的GluD1(蛋白)具有完全的NMDARs调控作用,而对AMPARs没有调控作用,相反地,含有细胞质序列基序4a的最短序列的GluD1完全调控AMPARs,而非NMDARs,但含有另一个序列基序4b的GluD1既不调控AMPARs也不调控NMDARs(图5d-i)。

图6 信号通路机制模型:NRXN1βSS4+-CBLN2-GluD1-NMDAR和NRXN3βSS4+-CBLN2-GluD1-AMPAR
(图片引自:Dai, J. et al., Nature 2021; 595: 261–265)
本研究表明,在海马突触中,NRXN1SS4+和NRXN3SS4+通过激活CBLN2复合物中的GluD1,特异性地增强NMDAR兴奋性突触后电流,抑制AMPAR兴奋性突触后电流。突触后GluD1将不同的突触前neurexins信号转导到不同的NMDAR和AMPAR应答中,即同源的突触前neurexins通过一个共同的效应体GluD差异地控制同源的突触后受体NMDAR和AMPAR,该效应体与被调节的受体具有相同的同源性。
研究也表明,由GluD1的短N端小脑肽结合域、CD4的跨膜区和GluD1的短C端序列基序组成的最小GluD1蛋白可以完全介导NMDARs和AMPARs的依赖于neurexins的调控。
此外,研究也发现,在突触前neurexins招募不同的突触后GluD复合物到突触后致部位,neurexins的特异性由不同的结合亲和力决定。 而且小脑肽和GluD2的表达是活性依赖的,表明这种信号机制也产生活性依赖的突触可塑性。
综上所述,这项研究揭示了一种意想不到的GluD1信号转导的分子机制,通过这种机制,neurexins信号控制AMPAR和NMDARs介导的突触回路应答(图6)。
原文连接:https://doi.org/10.1038/s41586-021-03661-6
【2】Neuron︱小胶质细胞C9orf72基因缺失促进突触丢失, 学习记忆缺陷的新机制
【3】Sci Adv︱周申如团队发现大脑脑区发育和脑区边界建立的新机制
【4】Nat Commun︱前沿! 宋源泉团队揭示抑制轴突再生的新分子机制
【5】Nat Neurosci︱MeCP2磷酸化: 快速抗抑郁药的持续抗抑郁作用的分子机制
【6】Mol Psychiatry︱前沿综述解读: 氯胺酮抗抑郁症的分子机制的最新进展
参考文献(上下滑动查看)
制版︱王思珍

